


Scientific journal
Fundamental research
ISSN 1812-7339
"Перечень" ВАК
ИФ РИНЦ = 1,798
EFFECT OF BACTERIA BACILLUS SUBTILIS ON THE LIPID PEROXIDATION OF SINAPIS ALBA UNDER CD-STRESS

В последние десятилетия в промышленно развитых странах увеличивается степень загрязнения окружающей среды тяжелыми металлами. Кадмий, относящийся к числу токсичных элементов, попадает в экосистемы, главным образом, в результате производства химических продуктов и использования фосфатных удобрений. Его концентрации в почвах угодий могут достигать высоких значений, а сам металл в виде ионов легко ассимилируется растениями [8]. Токсичность кадмия в растительном организме связана с увеличением перекисного окисления липидов (ПОЛ) и изменением антиоксидантной системы растений [5]. Следствием таких процессов могут быть нарушения метаболизма и гибель растений.
В настоящее время все большее значение для повышения устойчивости растений к стрессовым факторам приобретают исследования в области биотехнологий, использующих микроорганизмы. Ранее нами было показано протекторное действие клеток эндофитного штамма Bacillus subtilis 26Д на рост подсолнечника Helianthus annuus при действии ионов кадмия [3]. В связи с этим интересен вопрос о специфичности такого эффекта и механизмах его проявления, а также роли эндофитных бактерий в развитии окислительного стресса растений, вызванного тяжелыми металлами. Цель настоящей работы заключалась в изучении влияния обработки семян Sinapis alba клетками B. subtilis штаммов 26Д и 11ВМ на рост растений и образование малонового диальдегида (МДА) при стрессе, вызванном ионами кадмия.
Материалы и методы исследования
Объектом исследования служили растения горчицы белой (Sinapis alba L.). Семена перед посевом промывали мыльным раствором, затем выдерживали 1 мин в 96 %-м этаноле, ополаскивали в стерильной дистиллированной воде. В опытах использовали 20-часовую культуру бактерий, растущую на мясо-пептонном агаре при + 37 °С. Клетки бактерий отмывали 0,001 М KCl. Суспензию клеток доводили до необходимой концентрации по оптической плотности. 1 г семян обрабатывали в ламинар-боксе 20 мкл суспензии клеток бактерий с титром 1 млрд/мл.
Исследования проводили в чашках Петри и вегетационных сосудах в почве. Инокулированные и контрольные семена выращивали в чашках Петри (d = 140 мм, h = 24 мм) на фильтровальной бумаге, смоченной дистиллированной водой или раствором Cd(NO3)2•4H2O. Раствор соли готовили в пересчете на содержание иона металла. В вегетационных опытах использовали чернозем выщелоченный. В варианте с имитацией Cd-стресса после посева семян почву поливали раствором соли, а в контроле – дистиллированной водой. Растения выращивали при температуре 18–20 °С. Размещение вегетационных сосудов меняли каждый день по единой схеме, чтобы обеспечить более равномерную освещенность. В чашках Петри измерение сырой массы и концентрацию МДА проводили на 3, 4, 5 сутки от начала эксперимента; в вегетационных опытах – на 3, 6, 9, 14, 30 сутки.
Содержание МДА измеряли, используя метод Costa с соавторами [6], основанный на образовании окрашенного комплекса между МДА и тиобарбитуровой кислотой (ТБК) при нагревании. Концентрацию МДА определяли при 532 нм с помощью спектрофотометра UNICO 2800, вычитая величину неспецифической экстинкции при 600 нм. Содержание МДА рассчитывали с использованием коэффициента экстинкции, равного 155 мМ–1см–1. Все эксперименты проводили в трех биологических повторностях. В таблицах приведены средние арифметические значения и их стандартные ошибки.
Результаты исследования и их обсуждение
Интегральным показателем, характеризующим рост растений и его органов в условиях стресса, является биомасса. Сырая масса как побегов, так и корней растений, предобработанных клетками B. subtilis, была больше контрольных, что было особенно заметно на пятые сутки выращивания в чашках Петри (рисунок) и на тридцатые – в вегетационных опытах в почве (табл. 1). Так, обработка семян горчицы клетками обоих штаммов B. subtilis повышала массу корней в чашках Петри в среднем на 15 %, побегов – на 9 %. В вегетационных опытах масса побегов растений повышалась при инокуляции семян бациллами на 11 %.
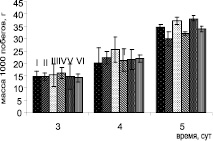 а
а 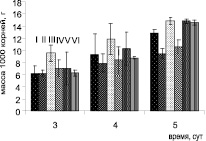 б
б
Влияние инокуляции семян бактериями B. subtilis на рост S. alba в присутствии ионов кадмия в чашках Петри: а – масса побегов; б – масса корней. Варианты опыта: I – контроль; II – растения, обработанные B. subtilis 26Д; III – растения, обработанные B. subtilis 11 ВМ; IV – Cd 20 мг/л; V – растения, обработанные B. subtilis 26Д + Cd 20 мг/л; VI – растения, обработанные B. subtilis11 ВМ + Cd 20 мг/л.
Таблица 1
Сырая масса побегов горчицы, выращенных в почве, загрязненной кадмием (100 растений, г)
|
Вариант |
Сутки |
|||||
|
3 |
6 |
9 |
14 |
30 |
||
|
Cd 0 мг/кг |
Контроль |
3,1 ± 0,5 |
6,0 ± 0,6 |
7,2 ± 0,6 |
8,7 ± 0,7 |
11,0 ± 0,3 |
|
B. subtilis 26Д |
3,2 ± 0,5 |
6,0 ± 0,5 |
7,2 ± 0,3 |
8,8 ± 0,8 |
12,4 ± 0,2 |
|
|
B. subtilis 11ВМ |
2,9 ± 0,3 |
5,9 ± 0,2 |
6,9 ± 0,3 |
8,8 ± 0,8 |
12,1 ± 0,2 |
|
|
Cd 10 мг/кг |
Контроль |
3,8 ± 0,9 |
5,4 ± 0,6 |
7,7 ± 0,6 |
8,1 ± 1,1 |
10,6 ± 1,1 |
|
B. subtilis 26Д |
2,8 ± 0,5 |
5,7 ± 0,4 |
8,2 ± 1,0 |
8,5 ± 1,1 |
12,4 ± 0,8 |
|
|
B. subtilis 11ВМ |
3,4 ± 0,6 |
5,9 ± 0,6 |
8,2 ± 0,3 |
9,4 ± 0,9 |
10,8 ± 0,7 |
|
|
Cd 200 мг/кг |
Контроль |
3,7 ± 0,6 |
6,3 ± 1,1 |
7,9 ± 0,9 |
9,2 ± 1,1 |
10,1 ± 0,8 |
|
B. subtilis 26 Д |
2,7 ± 0,3 |
6,0 ± 0,3 |
8,4 ± 0,3 |
9,3 ± 1,0 |
12,1 ± 0,5 |
|
|
B. subtilis 11ВМ |
3,6 ± 0,7 |
6,4 ± 0,7 |
9,3 ± 1,1 |
10,5 ± 0,9 |
11,9 ± 1,0 |
|
Ростстимулирующий эффект бактерий, вероятно, связан с тем, что бациллы могут продуцировать фитогормоно подобные вещества [1, 4], а также повышать содержание питательных веществ в доступной для растений форме [2] и подавлять развитие фитопатогенных микроорганизмов [4].
При выращивании горчицы в чашках Петри присутствие ионов кадмия в концентрации 20 мг/л вызывало снижение массы корней на пятые стуки на 34 %, а побегов – на 13 %. Два изученных штамма бацилл по-разному действовали на рост растений в присутствии ионов кадмия. Обработка семян штаммом B. subtilis 26Д не повышала устойчивость проростков к токсичным ионам металла – масса корней при Cd-стрессе также снижалась почти на 30 % в сравнении с инокулированными растениями, растущими на контрольной среде без кадмия. В отличие от штамма B. subtilis 26Д клетки штамма 11ВМ проявляли четкий протекторный эффект. Масса растений, предобработанных клетками бактерий B. subtilis 11ВМ, при Cd-стрессе не отличалась от таковой у контрольных растений, растущих на воде, что свидетельствует о защитном действии клеток этого штамма при действии токсичных ионов металла. Масса побегов растений, предобработанных клетками обоих штаммов бактерий, снижалась при действии кадмия так же, как и в контроле, в среднем на 13 %.
В вегетационных опытах при действии ионов кадмия в изученных концентрациях достоверно значимого протекторного эффекта выявить не удалось и можно говорить лишь о тенденции его проявления при обработке семян клетками бацилл.
Известно, что токсическое действие кадмия на растения проявляется в развитии окислительного стресса и образовании активных форм кислорода (АФК). АФК способны инициировать перекисное окисление липидов, в результате чего происходит повреждение мембранных структур. Кроме того, продукты ПОЛ (4-гидроксиалкенали, малоновый диальдегид и др.) обладают мутагенной активностью и блокируют клеточное деление [7].
Считается, что такой продукт ПОЛ, как малоновый диальдегид (МДА), может быть использован как биологический индикатор развития окислительного стресса растений при токсичном действии кадмия и других металлов [7].
При выращивании горчицы в чашках Петри достоверного влияния бацилл, а также ионов кадмия на содержание МДА в корнях растений выявить не удалось (табл. 2). При анализе этого показателя в побегах трехсуточных растений, предобработанных клетками бактерий, растущих на воде, отмечалась тенденция увеличения содержания МДА в сравнении с контролем. Затем эти различия сглаживались.
Таблица 2
Содержание МДА в растениях горчицы белой при воздействии ионов кадмия в водном растворе, 10–5 мМ/г сырого веса
|
Варианты |
Сутки |
|||
|
3 |
4 |
5 |
||
|
Корни |
||||
|
Cd 0 мг/л |
Контроль |
6,6 ± 1,3 |
6,7 ± 1,1 |
6,6 ± 0,4 |
|
B. subtilis 26Д |
6,4 ± 0,6 |
6,8 ± 0,6 |
6,2 ± 0,1 |
|
|
B. subtilis 11ВМ |
5,9 ± 0,8 |
6,1 ± 0,5 |
6,1 ± 0,4 |
|
|
Cd 20 мг/л |
Контроль |
5,8 ± 0,7 |
6,4 ± 1,5 |
6,1 ± 0,2 |
|
B. subtilis 26Д |
6,1 ± 0,9 |
7,2 ± 1,2 |
5,6 ± 0,4 |
|
|
B. subtilis 11ВМ |
5,8 ± 0,7 |
6,0 ± 0,6 |
5,8 ± 0,5 |
|
|
Побеги |
||||
|
Cd 0 мг/л |
Контроль |
10,7 ± 2,3 |
13,0 ± 1,9 |
12,09 ± 0,6 |
|
B. subtilis 26Д |
12,0 ± 2,7 |
11,0 ± 1,9 |
13,6 ± 0,9 |
|
|
B. subtilis 11ВМ |
14,8 ± 4,4 |
12,5 ± 2,3 |
11,5 ± 0,6 |
|
|
Cd 20 мг/л |
Контроль |
8,3 ± 1,0 |
9,8 ± 1,6 |
11,3 ± 0,6 |
|
B. subtilis 26Д |
10,6 ± 2,0 |
8,8 ± 1,5 |
9,9 ± 0,1 |
|
|
B. subtilis 11ВМ |
9,1 ± 1,6 |
11,2 ± 2,0 |
10,3 ± 0,4 |
|
При Cd-стрессе в побегах пятисуточных контрольных и предобработанных бактериями, растущих на воде, содержание МДА было выше, чем у растущих в присутствии ионов металла. При этом инокуляция проростков бациллами приводила к снижению уровня МДА в побегах при действии ионов кадмия.
В вегетационных опытах выявлено в среднем двукратное снижение уровня МДА в побегах контрольных растений при содержании ионов металла в концентрации 20 мг/кг и 200 мг/кг почвы. Вероятно, это связано с тем, что гумус черноземных почв способен адсорбировать ионы кадмия в таких концентрациях и, таким образом, не только снижать пресс Cd-стресса, но благоприятствовать реакциям растительных клеток, связанным со снижением ПОЛ.
Если в чашках Петри при инокуляции семян клетками бацилл мы смогли выявить лишь тенденцию повышения уровня МДА в сравнении с контрольными растениями, то в вегетационном опыте у трехсуточных растений наблюдали повышенный в 1,8 раза уровень МДА в сравнении с контрольными растениями (табл. 3). Это может быть связано с активным внедрением эндофитов в ткани растений горчицы в этом возрасте и ответной реакцией растительных клеток на этот процесс. Впоследствии в тканях побегов четырнадцати- и тридцатидневных растений, инокулированных бактериями, содержание МДА было ниже, чем у контрольных.
Таблица 3
Содержание МДА в побегах горчицы белой при воздействии ионов кадмия в почве (10–5 мМ/г сырого веса)
|
Вариант |
Сутки |
|||||
|
3 |
6 |
9 |
14 |
30 |
||
|
Cd 0 мг/кг |
Контроль |
6,4 ± 0,6 |
1,9 ± 0,3 |
2,9 ± 0,2 |
3,3 ± 0,2 |
3,5 ± 0,4 |
|
B. subtilis 26Д |
6,0 ± 0,6 |
1,3 ± 0,3 |
3,0 ± 0,1 |
2,1 ± 0,2 |
2,0 ± 0,2 |
|
|
B. subtilis 11ВМ |
5,9 ± 0,2 |
3,5 ± 0,1 |
2,5 ± 0,1 |
2,5 ± 0,1 |
2,2 ± 0,1 |
|
|
Cd 10 мг/кг |
Контроль |
3,1 ± 0,3 |
3,0 ± 0,1 |
3,2 ± 0,5 |
3,6 ± 0,2 |
4,3 ± 0,6 |
|
B. subtilis 26Д |
4,8 ± 0,9 |
2,8 ± 0,1 |
2,4 ± 0,2 |
2,9 ± 0,2 |
2,8 ± 0,1 |
|
|
B. subtilis 11ВМ |
5,6 ± 0,8 |
2,0 ± 0,1 |
1,4 ± 0,4 |
3,6 ± 0,7 |
3,1 ± 0,3 |
|
|
Cd 200 мг/кг |
Контроль |
3,0 ± 0,4 |
2,5 ± 0,4 |
2,4 ± 0,5 |
3,5 ± 0,6 |
5,5 ± 0,7 |
|
B. subtilis 26Д |
4,4 ± 0,2 |
1,1 ± 0,2 |
1,7 ± 0,2 |
2,3 ± 0,6 |
3,6 ± 0,2 |
|
|
B. subtilis 11ВМ |
5,3 ± 0,5 |
1,7 ± 0,1 |
1,5 ± 0,2 |
2,5 ± 0,4 |
3,7 ± 0,2 |
|
Аналогичная закономерность проявилась еще раньше – у девятисуточных растений при Cd-стрессе. В тканях побегов инокулированных бактериями растений пониженный в сравнении с контрольными уровень МДА при действии ионов кадмия сохранялся вплоть до тридцати суток (табл. 3).
Заключение
Таким образом, кроме известных ростстимулирующего эффекта эндофитных штаммов бактерий Bacillus subtilis по отношению к растениям, а также фунгицидного по отношению к фитопатогенным грибам [1, 4] нами выявлен протекторный эффект эндофитных бактерий при действии токсичных ионов кадмия. Анализ уровня МДА в тканях растений, указывающий на активность ПОЛ, выявил, что его содержание в тканях побегов растений горчицы, предобработанных клетками энофитов и растущих в почве в условиях Cd-стресса, ниже, чем в контрольных. Этот факт раскрывает один из механизмов защитного действия эндофитных штаммов бактерий на растения, который может быть связан со снижением интенсивности развития окислительного стресса и сохранением целостности мембранного аппарата растительных клеток, что важно для проявления их барьерной функции по отношению к ионам токсичного металла.
Рецензенты:
Карпов Д.Н., д.б.н., профессор, заведующий кафедрой биологии естественнонаучного факультета Стерлитамакского филиала Башкирского государственного университета, г. Стерлитамак;
Зейферт Д.В., д.б.н., доцент кафедры биологии естественнонаучного факультета Стерлитамакского филиала Башкирского государственного университета, г. Стерлитамак.
Работа поступила в редакцию 15.09.2014.
Библиографическая ссылка
Курамшина З.М., Смирнова Ю.В., Хайруллин Р.М. ВЛИЯНИЕ БАКТЕРИЙ BACILLUS SUBTILIS НА ПЕРЕКИСНОЕ ОКИСЛЕНИЕ ЛИПИДОВ SINAPIS ALBA ПРИ Cd-СТРЕССЕ // Фундаментальные исследования. 2014. № 9-11. С. 2460-2464;URL: https://fundamental-research.ru/en/article/view?id=35380 (дата обращения: 27.06.2026).