


Scientific journal
Fundamental research
ISSN 1812-7339
"Перечень" ВАК
ИФ РИНЦ = 1,798
EKOLOGO-TAKSONOMICHESKIY STRUCTURE OF THE MACROPHYTOBENTHOS OF THE AREA OF PRODUCTION OF PETROCONTAINING SHESKHARIS PNB SEWAGE (TSEMES BAY, THE BLACK SEA)

Прибрежная зона моря является первым уровнем, на который поступает основное количество загрязняющих веществ от источников, расположенных на его берегах. В этом плане, морские водоросли, являясь первичными продуцентами органического вещества, играют решающую роль в функционировании всей экосистемы водоёма и отдельных ее компонентов. Макрофитобентос выполняет роль мощного фильтра различных загрязняющих веществ, поступающих в его пределы, участвует в потреблении, распределении, переработке этих веществ и, по сути, обеспечении питания других структурных уровней шельфовой зоны моря.
Цемесская бухта по масштабам портово-промышленной и транспортной деятельности и, следовательно, по совокупности антропогенных воздействий на морскую среду не имеет себе равных в Азово-Черноморском регионе России. Существенный вклад в загрязнение прибрежной зоны восточного побережья бухты вносят нефтепродукты, попадающие в море в составе сточных вод перевалочной нефтебазы «Шесхарис» (далее по тексту – ПНБ «Шесхарис»). Начиная с 1963 и до 1977 г. сброс частично очищенных нефтесодержащих сточных вод предприятия осуществлялся из системы прудов-отстойников через подводный дюкер. В последующие 35 лет сточные воды отводятся в море в другом районе и только через глубоководный выпуск (удаление от берега 600 м). Расстояние от оголовка нового выпуска сточных вод предприятия до старого выпуска из прудов-отстойников составляет около 1 км.
Макрофиты, являясь прикрепленными организмами моря, представляют главную мишень воздействия нефтепродуктов, они особенно остро реагируют на любые изменения качества водной среды. Изучение видового состава и функционирования прибрежных водорослевых сообществ в условиях нефтяного загрязнения моря тесно связано с познанием процессов самоочищения водной среды и скорости восстановления фитоценозов.
Актуальность этой проблемы, исходя из требований российского законодательства в области охраны окружающей среды, а также с точки зрения решения практических задач гидротехнического и рекреационного строительства, очевидна.
Макрофитобентос Цемесской бухты относительно хорошо изучен. Большая часть работ посвящена изучению водорослей в летний период и в наиболее экологически напряженных районах (порт и прилегающая акватория). На сегодняшний день отсутствуют данные о современном состоянии макрофитобентоса в районе старого выпуска нефтесодержащих сточных вод ПНБ «Шесхарис» из системы прудов-отстойников, а по району нового глубоководного выпуска имеются лишь разрозненные данные разных лет исследований, выполненные в летний период года.
Цель данной работы – исследование эколого-таксономического разнообразия осеннего макрофитобентоса в районах старого и нового выпусков нефтесодержащих сточных вод ПНБ «Шесхарис».
Материал и методы исследований
Объектом исследований являлись водорослевые бентосные сообщества районов старого и нового выпусков нефтесодержащих сточных вод ПНБ «Шесхарис» (рис. 1).
Работы выполнялись на участке шельфа восточного побережья Цемесской бухты от нефтегавани до прудов-отстойников. Отбор качественных проб осуществлялся в конце сентября – начале октября 2013 г. по общепринятой гидроботанической методике (Калугина, 1969) с помощью водолаза с берега (глубины до 3 м) и малых плавсредств (глубины до 10 м). Всего было отобрано и обработано 32 качественные пробы водорослей. Видовой состав макрофитов определяли по «Определителю зеленых, бурых и красных водорослей южных морей СССР» [1] с учетом современных номенклатурных изменений [2–4]. Водоросли определены до вида. Для сравнительной оценки видовой структуры макрофитобентоса в районах наблюдений применены коэффициенты встречаемости (R), флористического сходства (КJ), флористический (Рфлор), предложенный Д.Т. Ченеем [7–9].
Результаты исследований и их обсуждение
На участке исследований прибрежная зона моря характеризуется выраженным микрорельефом дна с резко нарастающими глубинами. От уреза воды до глубины 5–7 м дно слагается галечниково-валунным материалом, пересечено скальными грядами, в складках – накопления мелкого гравийного материала с песком и признаками заиления. Глубже микрорельеф сглаживается, уступая место алеврито-пелитовым илам. На галечниках до глубины 0,3–0,5 м макроводоросли не обнаружены, что связано с высокой прибойностью и характером сложения донных грунтов.

Рис. 1. Карта-схема расположения районов исследований макрофитобентоса в Цемесской бухте
Проведенные исследования в двух районах (нового и старого выпусков) позволили обнаружить 57 видов макрофитов, относящихся к 33 родам, 21 семейству, 17 порядкам отделов Chlorophyta, Phaeophytа и Rhodophyta (таблица). По разнообразию таксономического состава водорослей доминируют виды отдела Rhodophyta, составляя более 57 % общего числа видов и родов, 48 % семейств, 41 % порядков. Наиболее многочисленный в видовом отношении порядок Ceramiales (3 семейства и 9 родов) включает 35,1 % общего числа видов макрофитобентоса. Основу видового разнообразия составляют семейства Ceramiaceae и Rhodomelaceae, на долю которых в сумме приходится 57,6 % общего числа видов данного отдела водорослей.
Видовой состав макрофитобентоса в районах выпусков нефтесодержащих сточных вод ПНБ «Шесхарис» (28.09. – 04.10.2013 г.)
|
№ п/п |
Отдел, вид |
Экологические показатели* |
Район |
|
|
Новый выпуск |
Старый выпуск |
|||
|
1 |
2 |
3 |
4 |
5 |
|
Phaeophyta |
||||
|
1. |
Myrionema balticum (Reinke) Foslie |
М |
– |
+ |
|
2. |
Ectocarpus confervoides (Roth) Le Jolis |
М |
– |
+ |
|
3. |
Entonema oligosporum (Strömf.) |
М |
+ |
+ |
|
4. |
Nereia filiformis (J.Ag.) Zanard. |
О |
– |
+ |
|
5. |
Sphacelaria cirrosa (Roth) Ag. |
М |
+ |
+ |
|
6. |
Cladostephus verticillatus (Lightf.) |
О |
+ |
+ |
|
7. |
Cystoseira barbata (Good. et Wood.) Ag. |
М |
+ |
+ |
|
8. |
C. crinita Bory |
О |
+ |
+ |
|
9. |
Padina pavonia (L.) Gaill. |
О |
+ |
– |
|
Chlorophyta |
||||
|
1. |
Acrosiphonia centralis (Lyngb.) Kjellm. |
О |
– |
+ |
|
2. |
Enteromorpha prolifera (O. Müll.) J. Ag. |
М |
+ |
+ |
|
3. |
E. linza (L.) J. Ag. |
М |
+ |
– |
|
4. |
E. intestinalis (L.) Link. |
П |
+ |
+ |
|
5. |
E. flexuosa (Wulf.) J. Ag. |
М |
+ |
– |
|
6. |
Chaetomorpha aёrea (Dillw.) Kütz. |
М |
+ |
+ |
|
7. |
Cladophora echinus (Bias.) Kütz. |
О |
– |
+ |
|
8. |
C. albida (Huds.) Kütz. |
П |
+ |
+ |
|
9. |
C. laetevirens (Dillw.) Kütz. |
П |
+ |
+ |
|
10. |
C. vadorum (Aresch.) Kütz. |
М |
+ |
+ |
|
11. |
C. liniformis Kütz. |
П |
+ |
– |
|
12. |
C. sericea (Huds.) Kütz. |
М |
– |
+ |
|
13. |
C. dalmatica Kütz. |
О |
– |
+ |
|
14. |
Cladophoropsis membranacea (Ag.) Börg. |
П |
+ |
+ |
|
15. |
Codium vermilara (Olivi) Delle Chiaje |
О |
+ |
+ |
|
Rhodophyta |
||||
|
1. |
Asterocytis ramosa (Thw.) Gobi |
П |
– |
+ |
|
2. |
Goniotrichum elegans (Chauv.) Zanard. |
М |
– |
+ |
|
3. |
Erythrotrichia carnea (Dillw.) J.Ag. |
П |
– |
+ |
|
4. |
Kylinia secundata (Lyngb.) Papenf. |
М |
+ |
+ |
|
5. |
K. virgatula (Harv.) Papenf. |
М |
+ |
– |
|
6. |
Gelidium crinale (Turn.) Lamour. |
О |
+ |
+ |
|
7. |
G. latifolium (Grev.) Born. et Thur. |
М |
+ |
+ |
|
8. |
Melobesia farinosa Lamour. |
? |
+ |
+ |
|
9. |
Corallina mediterranea Aresch. |
О |
+ |
+ |
|
10. |
C. granifera Ell. et Soland. |
О |
– |
+ |
|
11. |
Jania rubens (L.) Lamour. |
О |
+ |
– |
|
12. |
Gracilaria verrucosa (Huds.) Papenf. |
О |
– |
+ |
|
1 |
2 |
3 |
4 |
5 |
|
13. |
Phyllophora nervosa (DC.) Grev. |
О |
+ |
+ |
|
14. |
Antithamnion plumula (Ell.) Thur. |
П |
– |
+ |
|
15. |
Ceramium ciliatum (Ell.) Ducl. |
О |
+ |
+ |
|
16. |
C. rubrum (Huds.) Ag. |
П |
+ |
+ |
|
17. |
C. elegans Ducl. |
П |
+ |
+ |
|
18. |
C. tenuissimum (Lyngb.) J.Ag. |
М |
– |
+ |
|
19. |
C. diaphanum (Lightf.) Roth. |
П |
– |
+ |
|
20. |
Callithamnion corymbosum (J. E. Smith) Lyngb. |
М |
+ |
– |
|
21. |
Spermothamnion strictum (Ag.) Adriss. |
М |
+ |
+ |
|
22. |
Apoglossum ruscifolium (Turn.) J. Ag. |
М |
+ |
+ |
|
23. |
Polysiphonia denudata (Dillw.) Kütz. |
О |
+ |
+ |
|
24. |
P. subulifera (Ag.) Harv. |
М |
+ |
+ |
|
25. |
P. breviarticulata (Ag.) Zanard. |
? |
– |
+ |
|
26. |
P. opaca (Ag.) Zanard. |
М |
+ |
+ |
|
27. |
P. pulvinata Kütz. |
О |
– |
+ |
|
28. |
Lophosiphonia reptabunda (Suhr) Kylin |
М |
– |
+ |
|
29. |
Chondria tenuissima (Good. et Wood) Аg. |
О |
+ |
+ |
|
30. |
Ch. dasyphylla (Wood.) Ag. |
О |
– |
+ |
|
31. |
Laurencia paniculata J. Ag. |
О |
– |
+ |
|
32. |
L. obtusa (Huds.) Lamour. |
М |
+ |
+ |
|
33. |
L. hybrida (DC.) Lenorm. |
О |
– |
+ |
|
Всего |
36 |
50 |
||
Примечание. * – О – олигосапробы, М – мезосапробы, П – полисапробы; ? – не известно.
Количество видов Chlorophyta в 1,7 раза превышает Phaeophytа. Среди Chlorophyta по таксономическому составу доминирует порядок Cladophorales (60 % общего числа видов), у Phaeophytа распределение видов по отдельным надвидовым таксонам примерно одинаковое.
Большинство семейств (57,1 %) и родов (69,7 %) относятся к монотипическим. Высоким видовым разнообразием характеризуются Сladophoraceae, Rhodomelaceae и Ceramiaceae, на их долю приходится до 49 % общего числа зарегистрированных видов водорослей. Наибольшую видовую насыщенность имеет род Cladophora (7 видов), в составе родов Polysiphonia и Ceramium – по 5 видов. Среднее число видов в роде составляет 2,6.
Отличительной особенностью структуры фитоценозов исследованных районов является высокое видовое разнообразие (87,7 общего числа видов) сообществ у старого выпуска. Ведущее положение в структуре имеют Phaeophytа и Rhodophyta (составляют в сумме более 66 % видов). При этом в таксономической структуре Phaeophytа число родов и порядков не зависит от принадлежности сообщества к тому или иному исследованному району. Из Chlorophyta высоким видовым разнообразием отличается Сladophoraceae (66,7 % от общего числа видов отдела).
В районе нового выпуска зарегистрировано на 28 % видов меньше, чем у старого выпуска. Макроводоросли, принадлежащие к порядку Dictyotales, отмечены только в районе нового выпуска. Коэффициент флористического сходства состава выделенных таксонов альгофлор варьирует в интервале 32,7–35,7. По количеству видов наибольшее отличие обнаружено у Rhodophyta. Особенностью родовой структуры альгофлоры района нового выпуска является более низкая видовая насыщенность Polysiphonia, Ceramium и Cladophora и более высокая – Enteromorpha. Сопоставление изменчивости таксономической структуры отделов в каждом из характеризуемых районов показало, что наибольший предел вариаций числа видов прослеживается у порядков Rhodophyta и Chlorophyta, у семейств – Rhodomelaceae, Ceramiaceae и Сladophoraceae.
Малое количество бурых видов водорослей в исследованных районах (15,8 % общего числа видов), являющихся индикаторами чистых вод, определило высокое значение флористического коэффициента Чени (Рфлор = 5,3), что характерно для районов со средней степенью загрязнения морской воды (пределы Рфлор от 3 до 6). Распределение видов по группам сапробности показало превалирование олигосапробов в районе старого выпуска (38 % общего числа видов). Это позволяет сделать вывод, что основным фактором, влияющим на флористический состав макроводорослей в районе нового выпуска, является хроническое нефтяное загрязнение.
В целом макрофитобентос исследованных районов характеризуется выраженной индивидуальностью флористического состава (рис. 2). На это указывает и тот факт, что виды 4 порядков (Chordariales, Sporochnales, Goniotrichales, Bangeales) отмечены только у старого выпуска.
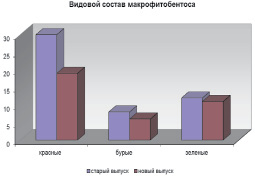
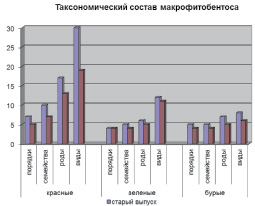
Рис. 2. Флористическая структура макрофитобентоса в районах старого и нового выпусков нефтесодержащих сточных вод ПНБ «Шесхарис»
Анализ распределения видов макроводорослей по величине коэффициента встречаемости показал, что в составе альгофлоры районов наибольшее количество видов (56,1 % общего числа видов) имеют низкий показатель (R = 1 – < 20 %) и относятся к I классу постоянства [5]. Доля видов, отнесенных к наивысшему V классу (R = 81–100 %), составляет всего 15,8 % общего числа видов.
Заключение
Район произрастания макроводорослей, а вернее уровень нефтяного загрязнения вод, определенным образом влияет на основные характеристики его водорослевых сообществ.
Альгофлора исследованных районов старого и нового выпусков нефтесодержащих сточных вод ПНБ «Шесхарис» насчитывает 57 видов макрофитов, относящихся к 33 родам, 21 семейству, 17 порядкам отделов Chlorophyta, Phaeophytа и Rhodophyta. По разнообразию таксономического и видового составов водорослей доминирует Rhodophyta, в его составе – порядок Ceramiales.
Таксономическая и видовая структура фитоценозов изученных районов имеет выраженные качественные различия. В составе макрофитобентоса районов отмечено около половины одинаковых видов. Сходство альгофлор подчеркивается доминированием количественных показателей отделов в последовательности Rhodophyta → Chlorophyta → Phaeophytа. Совпадение видовых таксонов увеличивается в ряду Phaeophytа → Chlorophyta → Rhodophyta.
В районе нового выпуска нефтесодержащих сточных вод таксономическая структура макрофитобентоса упрощенная, что обусловлено хроническим нефтяным загрязнением водной среды. У старого выпуска фитобентос находится в стадии естественных сукцессионных процессов, направленных на восстановление видового разнообразия и структуры сообществ.
Рецензенты:
Туркин В.А., д.т.н., профессор, начальник кафедры «Техносферная безопасность на транспорте», ФГБОУ ВПО «ГМУ им. адм. Ф.Ф. Ушакова», г. Новороссийск;
Страхова Н.А., д.т.н., профессор кафедры «Техносферная безопасность на транспорте», ФГБОУ ВПО «ГМУ им. адм. Ф.Ф. Ушакова», г. Новороссийск.
Библиографическая ссылка
Березенко Н.С. ЭКОЛОГО-ТАКСОНОМИЧЕСКИЙ СОСТАВ МАКРОФИТОБЕНТОСА РАЙОНА ВЫПУСКА НЕФТЕСОДЕРЖАЩИХ СТОЧНЫХ ВОД ПНБ «ШЕСХАРИС» (ЦЕМЕССКАЯ БУХТА, ЧЕРНОЕ МОРЕ) // Фундаментальные исследования. 2015. № 6-2. С. 219-224;URL: https://fundamental-research.ru/en/article/view?id=38544 (дата обращения: 07.07.2026).