


Scientific journal
Fundamental research
ISSN 1812-7339
"Перечень" ВАК
ИФ РИНЦ = 1,798
STRATEGY CHANGE OF GENTIANA PUNCTATA L. PARTIAL POPULATIONS IN UKRAINIAN CARPATHIANS

Исследование популяций редких видов растений не должно ограничиваться только рекомендациями по их охране и использованию. Первоочередной задачей остается углубленное изучение их стратегий в заповедных условиях, а также в условиях эксплуатации их ресурсов. Усилия в этом направлении должны концентрироваться на исследовании стратегии популяций редких видов и их адаптации к новым условиям среды. К группе таких таксонов принадлежит Gentiana punctata L. (Gentianaceae) – лекарственный краснокнижный вид [7]. Одной из причин исчезновения этого вида является заготовка местным населением его корневищ с целью использования в народной медицине. Определение стратегий популяций этого вида позволит разработать научно обоснованные подходы к сохранению и возобновлению его естественных запасов. Поэтому цель нашей работы – определить изменение стратегии частичных популяций G. punctata в разных условиях произрастания в Украинских Карпатах.
Материалы и методы исследования
Во время экспедиционных исследований, проведенных в 2009–2013 гг. в Украинских Карпатах, было изучено пять частичных популяций G. punctata, входящих в состав чорногорской (гора (г.) Брескул, г. Говерла), свидовецкой (г. Ворожеска, г. Татул) и мармарошской (г. Поп Иван) метапопуляционных организаций.
Для оценки межпопуляционной вариабельности сопоставляли морфометрические показатели репрезентативной выборки – по 50 генеративных особей с каждой популяции. Биометрический анализ данных проведен с помощью методов математической статистики [2]. Плотность, возрастную и пространственную структуры популяций исследовали на 20 пробных участках размером 1×1 м, заложенных методом случайных чисел [6]. Виталитетный анализ популяций проводили по методике Ю.А. Злобина (1989). По соотношению количества особей генеративного и вегетативного происхождения изучали характер самоподдержания.
Для выбора типа стратегии популяций использовали эколого-генетический подход, предложенный И. Цариком с соавт. [5], объединяющий методики Дж. Грайма [8-10] и О. Смирновой (1987) и учитывающий дифференциальные признаки на индивидуальном и групповом уровнях. Выделяли три типа первичных стратегий: конкурентный (К-тип), стресс-толерантный (S-тип), рудеральный (R-тип). Вторичные типы стратегий популяций определяли на основе комбинации дифференциальных признаков первичных типов [5].
Результаты исследования и их обсуждение
Изученные частичные популяции G. punctata различаются по условиям произрастания (антропогенное влияние, фитоценотическое окружение). Во всех субпопуляциях происходит неконтролируемое выкапывание корневищ местным населением, вытаптывание и повреждение генеративных побегов вблизи туристических маршрутов. Вытаптывание особей G. punctata на г. Татул связано с близким расположением частичной популяции к черничнику. Интенсивный выпас овец на г. Ворожеска приводит к отчуждению и вытаптыванию надземной части растений этого вида.
Биотическими факторами, негативно влияющими на частичные популяции, явлется преобладание плотнодерновинных злаков (г. Брескул, г. Говерла), Juniperus sibirica Burgsd. (г. Ворожеска) и Picea abies L. (г. Татул). Неблагоприятное влияние доминантных плотнодерновинных злаков (Poa sp, Nardus stricta L.) на горах Поп Иван и Татул компенсируется присутствием в сообществах низких кустарников (Vaccinium myrtillus L., Rhododendron kotschyi Simonk.) с высоким проективным покрытием, что препятствует формированию плотных дернин.
Согласно классификации О.В. Смирновой (1987), G. punctata принадлежит к растениям с длительным жизненным циклом и длительным прегенеративным периодом (5–10 лет) [3]. Особенности габитуса растений указывают на то, что G. punctata не в состоянии выдерживать конкуренцию с большинством доминантных видов высокогорных сообществ. Доказательством этого является также его неспособность к формированию популяций с высокой плотностью и диффузным расположением особей. Для G. punctata характерна метапопуляционная организация [5].
Условия произрастания растений определяют особенности протекания онтогенеза и способность популяций к самоподдержанию. Так, на г. Поп Иван и г. Татул наблюдается доминирование генеративного размножения над вегетативным (табл. 1). Это обусловлено неплотным задернением и наличием мест нарушения сплошного злакового покрова, что способствует прорастанию семян и приживлению подроста G. punctata. В условиях плотного задернения и интенсивного пасторального прессинга наблюдается уменьшение прорастания семян и приживления подроста, что и вызывает интенсификацию вегетативного размножения и его преобладание над генеративным на г. Брескул, г. Говерла, г. Ворожеска (табл. 1). Увеличение численности особей вегетативного происхождения рассматривается как усиление признаков стресс-толерантности [5].
В благоприятных эколого-географических условиях вегетативное размножение генеративных особей происходит за счет неглубокого омоложения к виргинильному состоянию (г. Поп Иван) (рисунок, а). Ухудшение условий роста и увеличение интенсивности антропогенного воздействия приводит к доминированию вегетативного размножения, к вегетативному размножению виргинильных особей, глубокому омоложению генеративных растений к виргинильному и имматурному состояниям (г. Брескул, г. Говерла, г. Татул, г. Ворожеска) (рисунок, б).
Несмотря на условия роста, во всех субпопуляциях G. punctata пик численности приходится на особи виргинильного состояния. Исследованные субпопуляции, кроме частичной популяции на г. Ворожеска, относятся к нормальным полночленным с левосторонним возрастным спектром (табл. 1). Отчуждение надземной и подземной массы населением при заготовлении лекарственного сырья и выпасе скота приводят к переходу субпопуляций от нормального к инвазионно-регрессивному или регрессивному типу, а также вызывает потерю дефинитивных признаков. Все вышеперечисленное указывает на усиление признаков стресс-толерантности [5].
Видовые особенности, тип самоподдержания, а также влияние фитоценотического окружения и антропогенной нагрузки предопределяют особенности пространственного размещения особей в субпопуляциях. Так, незначительное задернение почвы, доминирование генеративного размножения обусловливают диффузное расположение особей на г. Поп Иван (табл. 1). Ухудшение условий роста приводит к изменению пространственной структуры на компактно-диффузную (г. Говерла, г. Татул), а значительное задернение злаками и затенение кустарниками, интенсивный выпас скота вместе с преобладанием вегетативного размножения приводят к компактному расположению особей в субпопуляциях (г. Брескул, г. Ворожеска) (табл. 1). Это следует рассматривать как усиление признаков стресс-толерантности [5].
Таблица 1
Характеристики субпопуляций G. punctata в Украинских Карпатах
|
Характеристики |
Субпопуляции |
||||
|
г. Брескул |
г. Говерла |
г. Поп Иван |
г. Татул |
г. Ворожеска |
|
|
Местоположение |
1800–1900 S, N, E, W; 20–40° (> 45°) |
1930 S 35–45° |
1800–1850 NE 20–40° |
1650 N > 45° |
1735 SW 5–10° |
|
Плотность, ос/м2 |
0,6 |
3,1 |
7,2 |
1,4 |
0,3 |
|
Тип возрастной структуры |
Полночленная с левосторонним возрастным спектром |
Полночленная с левосторонним возрастным спектром |
Полночленная с левосторонним возрастным спектром |
Полночленная с левосторонним возрастным спектром |
Неполночленная, отсутствуют особи ювенильного состояния |
|
Пик числености особей (часть, %) |
Виргинильные (35,4 %) |
Виргинильные (26,1 %) |
Виргинильные (33,8 %) |
Виргинильные (41,7 %) |
Виргинильные (41,2 %) |
|
Тип пространственного размещения |
Компактное |
Компактно-диффузное |
Диффузное, на периферии компактно-диффузное |
Компактно-диффузное |
Компактное |
|
Генеративное / вегетативное размножение, % |
19,9 / 80,1 |
25,8 / 74,2 |
72,1 / 27,9 |
54,5 / 45,5 |
23,1 / 76,9 |
|
Виталитетный тип |
Процветающая |
Равновесная |
Равновесная |
Депрессивная |
Депрессивная |
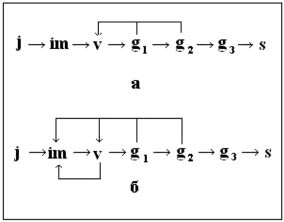
Последовательность прохождения возрастных состояний особями G. punctata в зависимости от условий произрастания и антропогенного воздействия (стрелками обозначены возможные пути переходов особей в разные возрастные состояния): а – в заповедных условиях; б – при воздействии неблагоприятных факторов
Высокая плотность особей наблюдается в популяции на г. Поп Иван, где отмечено неплотное задернение почвы и присутствие диких животных (табл. 1). Животные вызывают нарушение злакового задернения, что, в свою очередь, способствует прорастанию семян и приживлению подроста G. punctata. Высокая плотность растений в популяции является признаком конкурентности. Ухудшение условий произрастания и влияние антропогенного прессинга приводят к снижению плотности, в отдельных случаях до 0,3 ос/м2 (табл. 1), и усилению признаков стресс-толерантности.
Анализ морфометрических параметров растений свидетельствует о том, что самые высокие показатели генеративной и вегетативной сфер характерны для особей субпопуляции на г. Брескул (табл. 2), где зафиксирована низкая плотность и минимальное антропогенное воздействие. Морфометрические показатели растений на г. Говерла и г. Поп Иван несколько меньше, причина этого может заключаться во внутривидовой конкуренции за ресурсы питания в связи с высокой плотностью растений. В субпопуляциях на г. Татул и г. Ворожеска обнаружены самые малые показатели размеров и количества вегетативных и генеративных органов (табл. 2), что обусловлено влиянием неблагоприятных антропогенных и биотических факторов. Уменьшение количества и размеров органов надземной части способствует изменению виталитетной структуры с процветающей (г. Брескул) на равновесную (г. Говерла, г. Поп Иван) или депрессивную (г. Ворожеска, г. Татул) (табл. 1). Уменьшение морфометрических параметров растений является проявлением признаков стресс-толерантности [5].
Таблица 2
Морфометрические показатели генеративных особей G. punctata
|
Параметры |
Субпопуляции |
|||||
|
г. Брескул |
г. Говерла |
г. Поп Иван |
г. Татул |
г. Ворожеска |
||
|
1 |
2 |
3 |
4 |
5 |
6 |
|
|
Ширина листа, мм |
Х |
53 |
52 |
51 |
50 |
40 |
|
Sx |
±3,4 |
±4 |
±6 |
±6 |
±2 |
|
|
Sx’ |
13 |
14 |
18 |
19 |
7 |
|
|
СV, % |
24,5 |
26,9 |
35,3 |
38 |
17,5 |
|
|
Min |
25 |
29 |
25 |
23 |
25 |
|
|
Max |
55 |
75 |
80 |
76 |
50 |
|
|
Длина листа, мм |
Х |
183 |
125 |
118 |
114 |
127 |
|
Sx |
±12 |
±8 |
±11 |
±9 |
±7 |
|
|
Sx’ |
38 |
24 |
35 |
29 |
22 |
|
|
СV, % |
20,8 |
19,2 |
29,7 |
25,4 |
17,3 |
|
|
Min |
87 |
92 |
74 |
85 |
83 |
|
|
Max |
185 |
186 |
178 |
182 |
161 |
|
|
Площадь листа, мм2 |
Х |
654 |
471 |
387 |
338 |
321 |
|
Sx |
±69 |
±59 |
±44 |
±31 |
±31 |
|
|
Sx’ |
218 |
188 |
139 |
98 |
97 |
|
|
СV, % |
33,3 |
39,9 |
78,1 |
29,0 |
30,2 |
|
|
Min |
280 |
250 |
210 |
190 |
140 |
|
|
Max |
960 |
840 |
645 |
480 |
430 |
|
|
Число листьев на вегетативный побег, шт. |
Х |
5,9 |
5,9 |
5,5 |
5,1 |
4,8 |
|
Sx |
±0,4 |
±0,3 |
±0,4 |
±0,3 |
±0,3 |
|
|
Sx’ |
1,4 |
1,1 |
1,4 |
1,1 |
0,9 |
|
|
СV, % |
23,7 |
18,6 |
25,5 |
21,6 |
18,8 |
|
|
Min |
4 |
5 |
4 |
4 |
4 |
|
|
Max |
8 |
8 |
8 |
7 |
6 |
|
|
Диаметр вегетативного побега, мм |
Х |
83 |
75 |
69 |
59 |
50 |
|
Sx |
±0,5 |
±0,4 |
±0,4 |
±0,4 |
±0,3 |
|
|
Sx’ |
1,5 |
1 |
1 |
1 |
1 |
|
|
СV, % |
18,1 |
13,3 |
14,5 |
16,9 |
20 |
|
|
Min |
5 |
6 |
4 |
4 |
4 |
|
|
Max |
9 |
10 |
8 |
8 |
7 |
|
|
1 |
2 |
3 |
4 |
5 |
6 |
|
|
Число вегетативных побегов, шт. |
Х |
7,3 |
4,3 |
4,1 |
2 |
5,9 |
|
Sx |
±2,0 |
±0,7 |
±0,7 |
±0,3 |
±1,1 |
|
|
Sx’ |
6,5 |
2,2 |
2,3 |
0,8 |
3,4 |
|
|
СV, % |
89,0 |
51,2 |
56,1 |
40 |
57,6 |
|
|
Min |
1 |
1 |
1 |
1 |
1 |
|
|
Max |
19 |
8 |
8 |
3 |
13 |
|
|
Число генеративных побегов, шт. |
Х |
3,2 |
2,5 |
2,7 |
1,7 |
2,9 |
|
Sx |
±0,6 |
±0,5 |
±0,4 |
±0,2 |
±1,1 |
|
|
Sx’ |
2,0 |
1,5 |
1,3 |
0,7 |
1,4 |
|
|
СV, % |
62,5 |
60 |
48,1 |
41,2 |
48,3 |
|
|
Min |
1 |
1 |
1 |
1 |
1 |
|
|
Max |
8 |
6 |
5 |
3 |
12 |
|
|
Диаметр генеративного побега, мм |
Х |
9 |
6 |
7 |
7 |
6 |
|
Sx |
±0,9 |
±0,3 |
±0,4 |
±0,4 |
±0,2 |
|
|
Sx’ |
3 |
0,9 |
2 |
1 |
0,8 |
|
|
СV, % |
33,3 |
15 |
28,6 |
14,3 |
66,7 |
|
|
Min |
6 |
5 |
5 |
5 |
4 |
|
|
Max |
16 |
9 |
10 |
9 |
10 |
|
|
Высота генеративного побега, мм |
Х |
379 |
330 |
358 |
305 |
318 |
|
Sx |
±26 |
±23 |
±24 |
±12 |
±19 |
|
|
Sx’ |
83 |
72 |
74 |
37 |
60 |
|
|
СV, % |
21,9 |
21,8 |
20,7 |
12,1 |
18,9 |
|
|
Min |
260 |
225 |
270 |
259 |
243 |
|
|
Max |
495 |
435 |
495 |
350 |
420 |
|
|
Число метамеров генеративного побега, шт. |
Х |
5,1 |
4 |
4,3 |
4 |
4,1 |
|
Sx |
±0,3 |
±0,3 |
±0,3 |
±0,3 |
±0,3 |
|
|
Sx’ |
0,8 |
0,8 |
0,9 |
1,1 |
1 |
|
|
СV, % |
15,7 |
20 |
20,9 |
27,5 |
42,4 |
|
|
Min |
3 |
3 |
3 |
3 |
3 |
|
|
Max |
6 |
5 |
6 |
6 |
6 |
|
|
Длина метамеров генеративного побега, мм |
Х |
77 |
79 |
79 |
59 |
60 |
|
Sx |
±12 |
±13 |
±14 |
±7 |
±9 |
|
|
Sx’ |
38 |
41 |
42 |
28 |
29 |
|
|
СV, % |
49,4 |
51,9 |
53,1 |
47,5 |
48,3 |
|
|
Min |
30 |
18 |
25 |
19 |
25 |
|
|
Max |
150 |
140 |
140 |
102 |
103 |
|
|
Количество цветков в кольце, шт. |
Х |
4,8 |
4,2 |
4,9 |
3,7 |
3,6 |
|
Sx |
±0,4 |
±0,6 |
±0,7 |
±0,6 |
±0,5 |
|
|
Sx’ |
1,5 |
2 |
2,2 |
1,9 |
1,7 |
|
|
СV, % |
31,3 |
47,6 |
44,9 |
51,4 |
47,2 |
|
|
Min |
2 |
2 |
2 |
2 |
2 |
|
|
Max |
7 |
7 |
7 |
7 |
7 |
|
|
1 |
2 |
3 |
4 |
5 |
6 |
|
|
Количество цветков на побег, шт. |
Х |
8,6 |
8,5 |
8,6 |
7,4 |
7,2 |
|
Sx |
±0,2 |
±0,3 |
±0,3 |
±0,4 |
±0,3 |
|
|
Sx’ |
0,7 |
1,1 |
0,9 |
1,2 |
0,9 |
|
|
СV, % |
8,1 |
12,9 |
10,5 |
16,2 |
12,5 |
|
|
Min |
7 |
7 |
7 |
5 |
7 |
|
|
Max |
9 |
10 |
9 |
9 |
9 |
|
Примечание. Полужирным шрифтом выделены самые высокие морфометрические показатели.
Анализ репродуктивной способности популяций показал значительную зависимость генерирования особей G. punctatа от условий произрастания. Так, на г. Поп Иван в условиях неплотного задернения количество генеративных побегов составляло около 44 шт. на 10 м2. В то же время ухудшение фитоценотического окружения, проявляющееся в доминировании в сообществах плотнодерновинных и неплотнокустовых злаков, на г. Брескул и г. Говерла вызывает снижение репродуктивной способности. В указанных местопроизрастаниях количество генеративных побегов уменьшается до 22–28 шт./10 м2. Сочетание двух негативных воздействий, а именно затенения и задернения почвы и интенсивного антропогенного прессинга на г. Ворожеска и г. Татул приводит к существенному сокращению количества генеративных побегов (3–4 шт./10 м2). Вместе с сокращением количества генеративных побегов уменьшается и количество цветков в одном узле и на один генеративный побег (табл. 2). Так, в растений G. punctatа с гор Брескул, Говерла и Поп Иван насчитано по 8–9 цветков на один генеративный побег, а с гор Ворожеска и Татул – по 7–8.
Таким образом, субпопуляция на г. Поп Иван, подвергающаяся слабому стрессу и слабым нарушениям, характеризируется конкурентно-стресс-толерантными признаками (высокая плотность, левосторонний возрастной спектр, диффузная пространственная структура, большие морфометрические показатели, доминирование генеративного размножения и неглубокое вегетативное омоложение, высокие показатели репродуктивной способности). В условиях сильного стресса и слабых нарушений на г. Брескул и г. Говерла частичные популяции приобретают признаки стресс-толерантной стратегии (низкая плотность, преобладание виргинильных особей в возрастном спектре, доминирование вегетативного размножения, глубокое омоложение, компактно-диффузная или компактная пространственная структура). Их выживание обусловлено высокой репродуктивной способностью и большими морфометрическими параметрами. Субпопуляции, находящиеся в условиях сильного стресса и сильных нарушений (г. Татул, г. Ворожеска), являются стресс-толерантами на стадии угасания. Им свойственны следующие параметры: преобладание виргинильных особей в возрастном спектре, отсутствие ювенильных особей (г. Ворожеска), низкая плотность, доминирование вегетативного размножения (г. Ворожеска), компактная (г. Ворожеска) и компактно-диффузная (г. Татул) пространственная структура, глубокое вегетативное омоложение, малые морфометрические параметры, низкая репродуктивная способность, невозможность выдерживать межвидовую конкуренцию в фитоценозе.
Выводы
Таким образом, для G. punctata в Украинских Карпатах характерна стресс-толерантная стратегия: длительный онтогенез, невысокая плотность популяций, небольшие морфометрические параметры особей, неглубокое вегетативное омоложение, преобладание в возрастном спектре виргинильных особей, компактная и компактно-диффузная пространственная структура. Однако в благоприятных эколого-географических условиях и при незначительном затенении и задернении почвы в субпопуляциях этого вида проявляются признаки конкурентности, а интенсивное антропогенное воздействие, затенение высокими кустарниками и задернение почвы плотнодерновинными злаками приводят к их угасанию.
Рецензенты:Барна Н.Н., д.б.н., профессор кафедры ботаники и зоологии Тернопольского национального педагогического университета имени Владимира Гнатюка, г. Тернополь;
Иванников Р.В., д.б.н., старший научный сотрудник, Национальный ботанический сад им. Н.Н. Гришко Национальной академии наук Украины, г. Киев.
Работа поступила в редакцию 07.02.2014.
Библиографическая ссылка
Майорова О.Ю., Грицак Л.Р., Петрица В.А., Баран О.Н., Дробык Н.М. ИЗМЕНЕНИЕ СТРАТЕГИИ ЧАСТИЧНЫХ ПОПУЛЯЦИЙ GENTIANA PUNCTATA L. В УКРАИНСКИХ КАРПАТАХ // Фундаментальные исследования. 2014. № 3-2. С. 285-291;URL: https://fundamental-research.ru/en/article/view?id=33625 (дата обращения: 02.08.2026).