


Scientific journal
Fundamental research
ISSN 1812-7339
"Перечень" ВАК
ИФ РИНЦ = 1,798
THE FEATURES OF INTERPOPULATION VARIABILITY NEPETA GRANDIFLORA M. BIEB. INTRODUCED IN MOUNTAINOUS DAGHESTAN

Изучение закономерностей межпопуляционной дифференциации имеет большое значение для выявления ресурсного и адаптивного потенциала видов растений [4]. Эти закономерности проявляются при интродукции, поскольку выравнивание средовых условий позволяет оценить дифференциацию, которая является результатом предшествующей микроэволюции.
Популяционные исследования в настоящее время ведутся в различных направлениях. Среди различных подходов в изучении изменчивости растений особый интерес представляет анализ количественных морфологических признаков, так как именно они часто связаны с адаптивными свойствами организма [8].
Целью настоящей работы является изучение внутривидовой изменчивости и межпопуляционной дифференциации котовника крупноцветкового на основе комплекса морфологических признаков генеративного побега в эколого-географическом эксперименте.
Материалы и методы исследования
Котовник крупноцветковый (Nepeta grandiflora M. Bieb.) – многолетнее травянистое растение из семейства Lamiaceae. В Дагестане произрастает от низменности до субальпийского пояса – в лесах и ущельях, на лугах в высокотравье [7].
Материал для исследований был собран в 2010 г. в шести различных физико-географических районах Дагестана:
1. Окр. с. Алак Ботлихского района (1400 м над уровнем моря).
2. Окр. с. Ашар Курахского района (1650 м).
3. Гунибское плато, Гунибский р-он (1800 м).
4. Окр. с. Чираг Агульского района (2000 м).
5. Окр. с. Куруш Докузпаринского района (2200 м).
6. Агульский район, перевал Кокмадаг (2400 м).
Для сравнительного анализа адаптивных возможностей данного вида на Гунибскую экспериментальную базу (1750 м над уровнем моря) Горного ботанического сада ДНЦ РАН были пересажены по 30 растений из вышеназванных пунктов. Проводили учет прижившихся особей и фенологические наблюдения. Единицей учета в эксперименте является генеративный побег особи. Для изучения его структуры на второй год вегетации у 10 особей из каждой популяции срезали на уровне почвы по одному генеративному побегу
в фазе цветения.
Учитывали количество и длину междоузлий, количество боковых вегетативных и генеративных ветвей, длину, толщину и массу побега по фракциям (стебли, листья, соцветия) и массу 1000 семян. Определяли массу побега в целом, облиственность [6] и репродуктивное усилие [3]. Результаты измерений по каждой популяции были обработаны статистически с использованием программы Statistica 5.5. Уровни варьирования приняты по Зайцеву [2]: CV < 10 % – низкий, CV = 11–20 % – средний, CV > 20 % – высокий. Сила корреляции оценивалась по Доспехову [1]: r < 0,3 – слабая, r = 0,3–0,7 – средняя, r > 0,7 – высокая.
Результаты исследования и их обсуждение
Ход вегетации у интродуцированных популяций отражен в фенологических наблюдениях, приведенных в табл. 1. Фаза весеннего отрастания у них наступает в апреле с разницей от 5 до 13 дней. Обращают на себя внимание более поздние сроки отрастания интродуцентов, связанные с высотным уровнем произрастания исходных популяций. Это типичная адаптивная реакция, объясняемая предшествующим естественным отбором на максимальную приспособленность растений к конкретному местообитанию.
Таблица 1
Темпы прохождения фенофаз у популяций N. grandiflora
|
Происхождение популяции |
Высота над ур. моря, м |
Календарные даты начала фенофаз |
Кол-во дней от весеннего отрастания |
||||
|
отрастание |
стеблевание |
цветение |
плодоношение |
до цветения |
до плодоношения |
||
|
Ботлихский р-он, окр. с. Алак |
1400 |
09.04 |
24.05 |
13.07 |
17.08 |
95 |
130 |
|
Курахский р-он, окр. с. Ашар |
1650 |
14.04 |
24.05 |
15.07 |
19.08 |
92 |
127 |
|
Гунибский р-он, Гуниб. плато |
1800 |
17.04 |
24.05 |
18.07 |
24.08 |
92 |
129 |
|
Агульский р-он, окр. с. Чираг |
2000 |
22.04 |
20.05 |
16.07 |
20.08 |
85 |
123 |
|
Докузпаринский р-он, окр. с.Куруш |
2200 |
26.04 |
19.05 |
16.07 |
20.08 |
81 |
120 |
|
Агульский район, перевал |
2400 |
26.04 |
19.05 |
18.07 |
19.08 |
83 |
119 |
Вступление растений в генеративную фазу завершается к середине июля. Количество дней от весеннего отрастания до цветения для образцов из Ботлихского, Курахского и Гунибского районов составляет 92–95 дней в отличие от образцов из Агульского и Докузпаринского районов, у которых цветение наступает на одну-две недели раньше. Полный цикл развития от весеннего отрастания до плодоношения растения занимает 119–130 дней. Таким образом, факт укорачивания вегетационного периода у климатических экотипов с набором высоты в нашем эксперименте подтверждается.
Ростовые процессы у образцов также имеют свои особенности (рисунок). Наиболее интенсивно идут ростовые процессы до наступления фазы бутонизации, а к фазе цветения они замедляются. Прирост побегов по популяциям различается, но отрицательный тренд в динамике роста, т.е. чем ниже высотный уровень исходной популяции, тем больше прирост и, наоборот, выявляется на фазе цветения. Коэффициент корреляции (rxy) между высотным уровнем и высотой побега на этой фазе (июль) составлял –0,40.
Морфологическая характеристика генеративного побега N. grandiflora представлена в табл. 2. Как видно по данным таблицы, в условиях интродукции растения формируют значительную надземную массу, сильно ветвятся, хорошо облиственны в зоне ветвления. Длина побега варьируется в пределах 64,4–106,6 см и имеет тенденцию к уменьшению размеров побега, связанную с местом произрастания исходных популяций (высота над уровнем моря). Максимальное значение длины побега (106,6 см) отмечено для популяции из Ботлихского района (1400 м), далее идет уменьшение этого параметра до 64,4 см (Агульский район, 2400 м), исключение составляет популяция из Гунибского района (1800 м), для которой изменение условий среды произрастания незначительное. Уменьшение размеров годичного побега с набором высоты произрастания, наблюдаемое в условиях интродукции является, по всей видимости, результатом предшествующей микроэволюции.
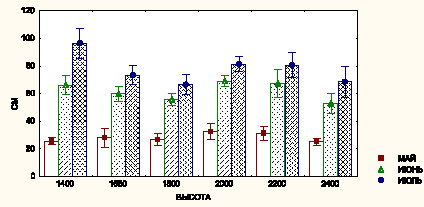
Темпы роста N. grandiflora в условиях интродукции (по оси абсцисс приведены высотные уровни исходных популяций, по оси ординат – высота побега)
Таблица 2
Характеристика морфологических признаков N. grandiflora в условиях интродукции
|
Пункты сбора |
Ботлихский р-он, 1400 м |
Курахский р-он, 1650 м |
Гунибский р-он, 1800 м |
Агульский р-он, с. Чираг, 2000 м |
Докузпаринский р-он, 2200 м |
Агульский р-он, перевал, 2400 м |
Объединенная выборка |
|||||||
|
|
CV, % |
|
CV, % |
|
CV, % |
|
CV, % |
|
CV, % |
|
CV, % |
|
CV, % |
|
|
Длина побега, см |
106,6 ± 3,12 |
9,25 |
72,2 ± 7,84 |
18,06 |
87,8 ± 3,53 |
12,73 |
75,4 ± 2,10 |
8,80 |
65,1 ± 1,98 |
9,60 |
64,4 ± 3,64 |
17,86 |
78,6 ± 2,21 |
21,74 |
|
Толщина стебля, мм |
6,9 ± 0,33 |
15,23 |
4,6 ± 0,16 |
10,68 |
4,6 ± 0,20 |
13,56 |
5,0 ± 0,18 |
11,19 |
4,4 ± 0,12 |
8,95 |
3,8 ± 0,15 |
12,41 |
4,9 ± 0,15 |
23,65 |
|
Кол-во междоузлий, шт. |
18,9 ± 0,10 |
1,67 |
19,0 ± 0,21 |
3,51 |
20,90 ± 0,28 |
4,19 |
17,7 ± 0,30 |
5,36 |
18,0 ± 0,30 |
5,24 |
17,70 ± 0,21 |
3,81 |
1,87 ± 0,17 |
7,19 |
|
Кол-во вегетативных ветвей, шт. |
9,6 ± 0,58 |
19,14 |
12,5 ± 1,21 |
30,46 |
6,9 ± 0,41 |
18,65 |
6,2 ± 0,44 |
22,56 |
7,1 ± 0,82 |
36,64 |
6,7 ± 0,56 |
26,37 |
8,2 ± 0,40 |
38,34 |
|
Кол-во генеративных ветвей, шт. |
4,2 ± 0,20 |
15,06 |
4,6 ± 0,27 |
18,33 |
7,0 ± 0,33 |
15,05 |
7,6 ± 0,45 |
18,81 |
5,5 ± 0,37 |
21,43 |
3,8 ± 0,29 |
24,18 |
5,5 ± 0,23 |
31,95 |
|
Длина вегетативной ветви, см |
18,4 ± 3,71 |
63,85 |
11,0 ± 0,83 |
23,81 |
14,8 ± 1,57 |
33,68 |
10,0 ± 1,36 |
42,93 |
10,2 ± 0,90 |
27,83 |
8,1 ± 0,97 |
37,88 |
12,1 ± 0,85 |
54,61 |
|
Длина генеративной ветви, см |
23,9 ± 3,13 |
41,37 |
10,7 ± 0,77 |
22,63 |
32,4 ± 2,52 |
24,56 |
18,4 ± 1,83 |
31,39 |
18,6 ± 1,49 |
25,30 |
16,1 ± 1,78 |
35,05 |
20,0 ± 1,19 |
46,21 |
|
Длина соцветия |
15,7 ± 0,88 |
17,73 |
10,8 ± 0,60 |
17,57 |
13,5 ± 0,94 |
22,05 |
17,6 ± 1,17 |
21,15 |
18,2 ± 1,51 |
26,15 |
10,6 ± 0,90 |
26,63 |
14,4 ± 0,56 |
30,34 |
|
Масса побега, г |
10,1 ± 0,99 |
31,12 |
5,2 ± 0,47 |
28,83 |
10,0 ± 0,87 |
27,55 |
6,1 ± 0,61 |
31,41 |
4,0 ± 0,31 |
24,87 |
3,0 ± 0,27 |
28,27 |
6,4 ± 0,44 |
53,17 |
|
Масса стеблей, г |
6,1 ± 0,65 |
33,85 |
2,8 ± 0,24 |
27,55 |
5,0 ± 0,58 |
36,61 |
2,9 ± 0,29 |
32,06 |
1,8 ± 0,13 |
21,74 |
1,6 ± 0,15 |
30,77 |
3,3 ± 0,26 |
60,80 |
|
Масса листьев, г |
3,7 ± 0,38 |
32,45 |
2,3 ± 0,24 |
33,15 |
3,8 ± 0,32 |
26,51 |
2,8 ± 0,28 |
31,86 |
1,7 ± 0,13 |
23,58 |
1,3 ± 0,15 |
37,12 |
2,6 ± 0,16 |
48,19 |
|
Масса соцветий, г |
0,3 ± 0,06 |
52,03 |
0,1 ± 0,02 |
40,67 |
1,2 ± 0,20 |
52,30 |
0,5 ± 0,07 |
48,03 |
0,4 ± 0,10 |
72,71 |
0,2 ± 0,05 |
85,56 |
0,5 ± 0,06 |
100,64 |
Как известно, длина побега складывается из количества междоузлий и их длины. При небольшом количестве междоузлий, характерном для выборки из Ботлихского района, высокорослость побегов этой популяции достигается, по всей видимости, за счет длины самих междоузлий. Средняя длина междоузлия данного образца максимальная по сравнению с другими и составляет 5,6 см. Характер изменчивости длины междоузлий для изученных популяций имеет свои особенности. Длина междоузлий побегов Агульской популяции (перевал) с 1-го по 2-й увеличивается, с 3-го по 5-й на одном уровне, с 6-го по 10-й уменьшается, затем увеличивается до предпоследнего. Для Курахской популяции изменение длины междоузлий происходит несколько по-другому. Со 2-го по 16-й длина междоузлий то уменьшается, то увеличивается. Что касается выборок из Ботлихского, Гунибского и Агульского (Чираг) районов, то у них с 1-го по 3-й данный параметр увеличивается, начиная с 4-го по 8-й уменьшается, затем идет увеличение до предпоследнего междоузлия. У Докузпаринской популяции идет нарастание длины междоузлий в начале вегетативной части побега, а в генеративной – происходит уменьшение. По-видимому, в условиях интродукции, при выравнивании средовых факторов, различие в изменчивости длины междоузлий, приобретенное в процессе естественного отбора в природных условиях, сохраняется.
Сухая масса годичного побега и число побегов на особь характеризуют мощность роста и развития особи [5]. Результаты оценки накопления сухой массы побега показывают (табл. 2), что она изменчива у популяций и имеет размах от 3,0 до 10,1 г, при этом максимальное значение массы отмечено у выборок из Ботлихского и Гунибского районов. Масса побегов формируется за счет длины и толщины стебля, количества листьев, боковых ветвей и их длины. В первой декаде июня на генеративном побеге формируются боковые вегетативные и генеративные ветви. У Ботлихской, Курахской и Гунибской популяций (табл. 3) максимальную длину вегетативные ветви имеют в 6–7 узлах; у других популяций в 4–5-м. Что касается генеративных ветвей, то у первых трех популяций максимальная длина их в 11–14-м узле, у остальных в 7–8 узлах. В Ботлихской и Курахской популяциях количество боковых вегетативных ветвей более чем в два раза превосходит число генеративных, а в Гунибской и Агульской (2000 м) превалирует число генеративных, тогда как в объединенной выборке вегетативных ветвей в 1,5 раза больше генеративных. Длина как вегетативных, так и генеративных ветвей различается по популяциям. Бóльшая длина ветвей, естественно, отмечена в популяциях, где высокие значения массы побега (Ботлихская и Гунибская). Но во всех популяциях значения длины генеративных ветвей выше, чем вегетативных.
Таблица 3
Характеристика боковых ветвей генеративного побега N. grandiflora
|
Признаки
Популяции |
Номер узла |
Максимальная длина |
||
|
вег. ветви |
ген. ветви |
вег. ветви, см |
ген. ветви, см |
|
|
Ботлихский р-он, окр. с. Алак |
7 |
12 |
10,8 |
13,0 |
|
Курахский р-он, окр. с. Ашар |
6 |
14 |
10,7 |
14,8 |
|
Гунибский р-он, Гуниб. плато |
6 |
11 |
8,5 |
12,0 |
|
Агульский р-он, окр. с. Чираг |
4 |
10 |
7,5 |
11,6 |
|
Докузпаринский р-он, окр. с. Куруш |
5 |
7 |
7,2 |
9,6 |
|
Агульский район, перевал |
5 |
8 |
7,8 |
9,1 |
Значения толщины стебля варьируются в пределах 3,8–6,9 мм и меняются соразмерно длине побега; толщина стебля максимальна при максимальной длине побега (Ботлихская популяция – 6,9 и 106,6 см) и минимальна при минимальной (Агульская (2400 м – 3,8 и 64,6).
При сравнительном анализе изменчивости учтенных признаков обнаружено, что весовые признаки всех популяций находятся на высоком уровне изменчивости. Количество междоузлий, толщина стебля и длина соцветий имеют, соответственно, низкий и средний уровень изменчивости. Наиболее вариабельна такие признаки, как число и длина вегетативных и генеративных ветвей.
Структура распределения сухой массы побега по фракциям показывает, что она носит неодинаковый характер. Масса стеблей во всех популяциях выше массы листьев. Но в Ботлихской и Гунибской популяциях эта разница значительна (3,7 и 3,8 против 6,1 и 5,0 соответственно). Масса соцветий имеет минимальную долю от общей во всех популяциях, но в Гунибской популяции она значительна – 1,2 г, тогда, как в остальных колеблется в пределах 0,1–0,5 г.
Определение репродуктивного усилия (табл. 4) у изученных популяций показывает, что оно колеблется в пределах 2,7–11,8 %. Важно подчеркнуть, что репродуктивное усилие не всегда связано с продуктивностью растений. Так, при средней массе побега в Гунибской популяции, равной 10,0 г, репродуктивное усилие составляет 11,8 %, тогда как при массе побега в Ботлихской популяции 10,1 г репродуктивное усилие составляет всего 3,4 %.
Репродуктивный успех популяций в новых условиях зависит, по всей видимости, от степени различия с исходными местообитаниями. Различие это в Гунибской популяции незначительно, чем и объясняется здесь высокое значение Re.
Помимо общей продуктивности большой интерес представляют данные по облиственности побегов (табл. 4). По полученным данным, низкая облиственность наблюдается в Ботлихской и Гунибской популяциях при максимальной среди
изученных популяций средней массе побега 10,1 и 10,0 г соответственно. В данном случае, очевидно, масса побега достигла максимального значения за счет стеблевой. В остальных четырех популяциях доли стеблевой и листовой массы сравнительно приближены друг к другу. В целом по популяциям облиственность не находится в положительной коррелятивной связи с продуктивностью побега.
Таблица 4
Репродуктивное усилие и облиственность побегов N. grandiflora
|
Происхождение популяции |
Высота над уровнем моря, м |
Облиственность, % |
Репродуктивное усилие (Re), % |
|
Ботлихский р-он, окр. с. Алак |
1400 |
36,5 |
3,5 |
|
Курахский р-он, окр. с. Ашар |
1650 |
43,5 |
2,7 |
|
Гунибский р-он, Гуниб. плато |
1800 |
38,3 |
11,8 |
|
Агульский р-он, окр. с. Чираг |
2000 |
44,7 |
7,9 |
|
Докузпаринский р-он, окр. с. Куруш |
2200 |
43,4 |
10,4 |
|
Агульский район, перевал |
2400 |
44,3 |
5,7 |
Изменчивость побегов различных популяций по признакам генеративной сферы имеет определенный интерес с точки зрения микроэволюции. Основные показатели изменчивости массы 1000 семян представлены в табл. 5. Согласно полученным данным, средние значения этого весового признака колеблются в пределах 0,537–0,821 г. Масса семян минимальная для популяции из Курахского района (1650 м) и максимальная для популяции из Ботлихского района (1400 м).
Таблица 5
Сравнительная характеристика интродуцированых популяций
N. grandiflora по массе 1000 семян
|
Происхождение популяции |
Высота над уровнем моря, м |
Масса семян |
|
|
|
CV, % |
||
|
Ботлихский р-он, окр. с. Алак |
1400 |
0,821 ± 0,005 |
7,3 |
|
Курахский р-он, окр. с. Ашар |
1650 |
0,511 ± 0,008 |
16,9 |
|
Гунибский р-он, Гуниб. плато |
1800 |
0,569 ± 0,012 |
13,2 |
|
Агульский р-он, окр. с. Чираг |
2000 |
0,537 ± 0,031 |
8,6 |
|
Докузпаринский р-он, окр. с. Куруш |
2200 |
0,539 ± 0,019 |
16,5 |
|
Агульский район, перевал |
2400 |
0,567 ± 0,073 |
9,3 |
По коэффициенту вариации масса семян по популяциям находится на среднем и низком уровне изменчивости. Выявленные в условиях интродукции межпопуляционные различия по массе семян, вероятно, можно рассматривать как результат приспособления к условиям интродукции.
Корреляционный анализ, проведенный по морфологическим признакам генеративного побега, показал наличие положительной корреляционной связи по большинству пар учтенных признаков (табл. 6). Слабые корреляционные связи отмечены между признаками: длина побега – количество вегетативных и генеративных ветвей, длина и масса соцветий; толщина стебля – количество междоузлий и вегетативных ветвей, длина и масса соцветий; количество междоузлий – количество вегетативных ветвей и длина соцветий; масса побега и масса стеблей – количеством вегетативных и количеством генеративных ветвей, длиной соцветий. Количество вегетативных ветвей находится в отрицательной корреляции с количеством и длиной генеративных ветвей, длиной и массой соцветий. Для остальных пар признаков связь оценена как средняя и сильная.
Однофакторный дисперсионный анализ выявил степень влияния высотного уровня исходных популяций на изменчивость морфологических признаков генеративного побега N. grandiflora в условиях интродукции (табл. 7). Результаты свидетельствуют о существенном (на высоком уровне значимости) влиянии этого фактора на изменчивость всех учтенных, за исключением одного – длина вегетативных ветвей, морфологических признаков побега. Причем сила влияния фактора выше на такие признаки, как длина побега и толщина стебля. При регрессионном анализе выяснилось, что достаточно значимо влияние высотного фактора на признаки вегетативной сферы (длина побега и толщина стебля), где коэффициент детерминации составляет соответственно 52,90 и 48,45 %. Однако влияние этого фактора на все признаки генеративной сферы незначительное, т.е. изменчивость их носит случайный характер. Почти все изученные признаки находятся в отрицательной корреляционной связи с высотой происхождения исходных популяций на высоком уровне значимости (значимости (p ≤ 0,001).
Таблица 6
Корреляционные связи между признаками генеративного побега в объединенной выборке интродуцированных популяций N. grandiflora
|
Признаки |
Длина побега |
Толщина стебля |
Кол-во междоузлий |
Кол-во вегет. ветвей |
Кол-во генер. ветвей |
Длина вегет. ветви |
Длина генер. ветви |
Длина соцветия |
Масса побега |
Масса стеблей |
Масса листьев |
Масса соцветий |
|
Длина побега |
1,00 |
0,77*** |
0,53*** |
0,15 |
0,07 |
0,40** |
0,51*** |
0,22 |
0,82*** |
0,87*** |
0,74*** |
0,29* |
|
Толщина стебля |
0,77*** |
1,00 |
0,23 |
0,28* |
–0 |
0,33** |
0,36** |
0,29* |
0,71*** |
0,77*** |
0,66*** |
0,02 |
|
Кол-во междоузлий |
0,53*** |
0,23 |
1,00 |
0,14 |
0,33** |
0,30* |
0,57*** |
0,03 |
0,61*** |
0,56*** |
0,57*** |
0,60*** |
|
Кол-во вегетативных ветвей |
0,15 |
0,28* |
0,14 |
1,00 |
–0,34** |
0,15 |
–0,31* |
–0,31* |
0,11 |
0,16 |
0,17 |
-0,34** |
|
Кол-во генеративных ветвей |
0,07 |
–0 |
0,33** |
–0,34** |
1,00 |
0,01 |
0,38** |
0,44*** |
0,29* |
0,15 |
0,34** |
0,52*** |
|
Длина вегетативной ветви |
0,40** |
0,33** |
0,30* |
0,15 |
0,01 |
1,00 |
0,32* |
0,15 |
0,43** |
0,43*** |
0,36** |
0,22 |
|
Длина генеративной ветви |
0,51*** |
0,36** |
0,57*** |
–0,31* |
0,38** |
0,32* |
1,00 |
0,34** |
0,72*** |
0,66*** |
0,61*** |
0,74*** |
|
Длина соцветия |
0,22 |
0,29* |
0,03 |
–0,31* |
0,44*** |
0,15 |
0,34** |
1,00 |
0,22 |
0,16 |
0,20 |
0,36** |
|
Масса побега |
0,82*** |
0,71*** |
0,61*** |
0,11 |
0,29* |
0,43*** |
0,72*** |
0,22 |
1,00 |
0,97*** |
0,94*** |
0,49*** |
|
Масса стеблей |
0,87*** |
0,77*** |
0,56*** |
0,16 |
0,15 |
0,43*** |
0,66*** |
0,16 |
0,97*** |
1,00 |
0,88*** |
0,36** |
|
Масса листьев |
0,74*** |
0,66*** |
0,57*** |
0,17 |
0,34** |
0,36** |
0,61*** |
0,20 |
0,94*** |
0,88*** |
1,00 |
0,39** |
|
Масса соцветий |
0,29* |
0,02 |
0,60*** |
–0,34** |
0,52*** |
0,22 |
0,74*** |
0,36** |
0,49*** |
0,36** |
0,39** |
1,00 |
Таблица 7
Результаты дисперсионного и регрессионного анализов изменчивости морфологических признаков интродуцированных популяций N. grandiflora
|
Признаки |
Компоненты дисперсии |
rxy |
|
|
h2 |
r2 |
||
|
Длина побега, см |
75,39*** |
52,90*** |
–0,73*** |
|
Толщина стебля, мм |
72,24*** |
48,45*** |
–0,70*** |
|
Кол-во междоузлий, шт. |
69,98*** |
20,27*** |
–0,45*** |
|
Кол-во вегет. ветвей, шт. |
51,17*** |
21,99*** |
–0,47*** |
|
Кол-во генер. ветвей, шт. |
67,29*** |
0,06 |
0,03 |
|
Длина вегет. ветвей, см |
27,99** |
20,28*** |
–0,45*** |
|
Длина генер. ветвей, см |
54,66*** |
3,48 |
–0,19 |
|
Длина соцветий, см |
48,24*** |
0,01 |
–0,01 |
|
Масса побега, г |
66,90*** |
41,20*** |
–0,64*** |
|
Масса стеблей, г |
65,57*** |
46,79*** |
–0,68*** |
|
Масса листьев, г |
58,96*** |
36,05*** |
–0,60*** |
|
Масса соцветий, г |
57,72*** |
0,83 |
–0,09 |
Выводы
Проведенные исследования котовника крупноцветкового в условиях интродукции позволяют сделать следующие выводы:
1. Наблюдаемое укорачивание вегетационного периода в интродукционных условиях относительно набора высоты произрастания исходных популяций является типичной адаптивной реакцией, объясняемой предшествующим естественным отбором на максимальную приспособленность растений к конкретному
местообитанию.
2. Изменчивость морфологических признаков в условиях интродукции не подчинена какой-либо закономерности. У изученных популяций наблюдается сходство по уровню изменчивости весовых признаков. Большая часть изученных признаков находится в положительной корреляционной связи между собой и отрицательной с высотным происхождением популяций.
3. Полученные данные имеют интерес для понимания механизмов приспособительных реакций интродуцентов и могут найти применение для интродукционного прогнозирования.
Рецензенты:Магомедмирзаев М.М., д.б.н., главный научный сотрудник, ФГБУН «Горный ботанический сад ДНЦ РАН», г. Махачкала;
Магомедова М.А., д.б.н., профессор, зав. кафедрой ботаники, ФГБОУ «Дагестанский государственный университет», г. Махачкала.
Работа поступила в редакцию 31.01.2014.
Библиографическая ссылка
Курамагомедов М.К., Гусейнова З.А. ОСОБЕННОСТИ МЕЖПОПУЛЯЦИОННОЙ ИЗМЕНЧИВОСТИ NEPETA GRANDIFLORA M. BIEB. ПРИ ИНТРОДУКЦИИ В ГОРНЫХ УСЛОВИЯХ ДАГЕСТАНА // Фундаментальные исследования. 2014. № 3-1. С. 93-99;URL: https://fundamental-research.ru/en/article/view?id=33592 (дата обращения: 04.07.2026).