


Scientific journal
Fundamental research
ISSN 1812-7339
"Перечень" ВАК
ИФ РИНЦ = 1,798
THE INFRADIAN RHYTHM OF LOCOMOTOR ACTIVITY AT MALE CHAFFINCH (FRINGILLA COELEBS) DURING AUTUMN MIGRATION

Волнообразность числа пролетающих птиц была отмечена В.Р. Дольником при ежедневной регистрации осенних миграций на Куршской косе и в пяти пунктах Прибалтики [3]. Далеко не все регистрируемые максимумы числа пролетающих птиц можно было связать с динамикой погодных условий. Автором указывается на совпадение календарных дат миграционных волн в разные годы. В период наблюдений с 1958 по 1962 гг. на Куршской косе максимальное число мигрирующих птиц зарегистрировано: 17–18, 21–22, 25–27, 29–30 сентября, 3–4, 7–8 октября [4]. Однако практически нет работ по изучению многодневных (инфрадианных) ритмов локомоторной активности птиц в сезон миграции.
Целью работы было исследование инфрадианных ритмов уровня локомоторной активности самцов зябликов в период осенней миграции.
Материалы и методы исследования
Эксперимент проведен на самцах зябликов (Fringilla coelebs). Взрослые особи (n = 6) были пойманы весной, а молодые (n = 8) – в июле, в птенцовом (ювенальном) оперении. При работе с экспериментальными животными руководствовались приказом Минздрава СССР № 755 от 12.08.1977 г. Локомоторную активность птиц, отражающую миграционное беспокойство, как и другие исследователи, оценивали по числу прыжков на жердочку за сутки [3]. Птиц содержали в отдельных клетках 40×25×30 с двумя жердочками. Один конец жердочки крепился неподвижно и имел на себе противовес, на другом подвижном конце имеющем ограничитель сверху и снизу располагался счетчик (Sigma-500, Германия), регистрирующий отклонение этого конца жердочки при прыжке птицы. Исследование проводили в двух разных комнатах: в одной поддерживалось искусственное освещение полного спектра (5600 К) в 2200 лк на уровне жердочек с режимом освещения, имитирующим естественное для этого сезона, в другой было естественное освещение. Температура в обеих комнатах поддерживалась в пределах +20–23 °С. В кормушках всегда присутствовали семена рапса и разрезанного пополам подсолнечника. Показатели счетчика регистрировали после прекращения птицами дневной активности ежедневно с 14 сентября по 28 октября 2011 г. Всего было проведено за 45 дней 630 измерений.
Статистическую обработку полученных результатов проводили с использованием программы «Statistica 6.0». Достоверность различий между показателями определяли по U-критерию Манна‒Уитни. Различия считали статистически значимыми при p < 0,05.
Результаты исследования и их обсуждение
В динамике показателей локомоторной активности у 4-х птиц, содержавшихся в условиях естественного освещения, а также у 7 птиц в условиях искусственного освещения был выявлен 8-суточный биоритм. У трех взрослых птиц, не завершивших линьку к 15 сентября, локомоторная активность была на низком уровне и достоверных ритмических колебаний этого показателя не наблюдалось.
Выявленный 8-суточный биоритм локомоторной активности у птиц, содержавшихся в условиях естественного освещения, был синхронен у всех особей, за исключением одного (№ 1) (рис. 1). У этого зяблика после 10-го октября наблюдалось смещение фазы 8-суточного ритма локомоторной активности на 4 сут.
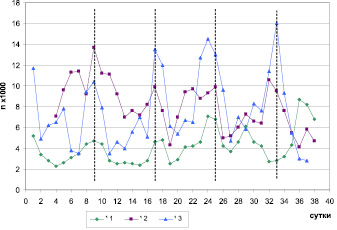
Рис. 1. Динамика уровня локомоторной активности трех самцов зябликов (№ 1 – ad; № 2 и № 3 – sad) в период с 14 сентября по 21 октября 2011 г., содержавшихся в условиях естественного освещения. По оси абсцисс – порядковый номер суток от начала исследования. По оси ординат – число прыжков за сутки
У птиц, содержавшихся в условиях искусственного освещения, 8-суточный ритм локомоторной активности либо совпадал по фазе, либо находился в противофазе по отношению к фазе этого ритма в группе зябликов, находившихся при естественном освещении. Таким образом, относительно фазы 8-суточного ритма локомоторной активности зябликов, находившихся при естественном освещении, птицы, содержавшиеся в условиях искусственного освещения, разделились на 2 группы: совпадающие по фазе – 1-я группа (n = 4) и находящиеся в противофазе – 2-я группа (n = 3).
С целью выявления статистической значимости различий показателей в акрофазе и батифазе у птиц, содержавшихся в условиях искусственного освещения, был применен метод наложения эпох. Ни одна птица не была исключена из анализа по причине отклонения фазы ритма локомоторной активности. Весь исследуемый период был разбит на 8-суточные периоды. Значения процентного отклонения от индивидуального среднего уровня локомоторной активности за 14, 22, 30 сентября, 8 и 16 октября были отнесены к первому дню 8-суточного периода; 15, 23 сентября, 1, 9 и 17 октября – ко второму дню 8-суточного периода; 16, 24 сентября, 2, 10 и 18 октября – к третьему дню 8-суточного периода; 17, 25 сентября, 3, 11 и 19 октября – к четвертому дню 8-суточного периода; 18, 26 сентября, 4, 12 и 20 октября – к пятому дню 8-суточного периода; 19, 27 сентября, 5, 13 и 21 октября – к шестому дню 8-суточного периода; 20, 28 сентября, 6 и 14 октября – к седьмому дню 8-суточного периода; 21, 29 сентября, 7 и 15 октября – к восьмому дню 8-суточного периода (рис. 2).
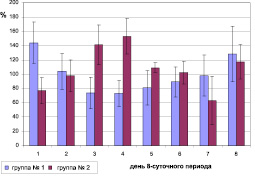
Рис. 2. Распределение локомоторной активности по дням 8-суточного периода у двух групп птиц, содержащихся в условиях искусственного освещения (группа № 1 – биоритм локомоторной активности совпадает по фазе с птицами, содержавшимися при естественном освещении, группа № 2 – биоритм локомоторной активности которых был в противофазе с птицами, содержавшимися при естественном освещении). По оси абсцисс – день 8-суточного периода. По оси ординат – % от индивидуального среднего уровня локомоторной активности
Использованный метод наложения эпох позволил установить, что у 1-й группы зябликов из числа птиц, содержавшихся в условиях искусственного освещения, значения уровня локомоторной активности в акрофазе (максимальные значения) приходились на 1-й день этого 8-суточного периода и статистически значимо различались со значениями в батифазе (минимальные значения – 5-й день 8-суточного периода). Для 2-й группы, наоборот, акрофаза уровня локомоторной активности приходилась на 5-й день 8-суточного периода и статистически значимо различалась с ее значениями в батифазе (1-й день 8-суточного периода). Таким образом, все максимумы локомоторной активности приходились либо на 14, 22, 30 сентября, 8 и 16 октября (первая группа), либо на 18, 26 сентября, 4, 12 и 20 октября (вторая группа).
У трех птиц (две молодые при естественном и одна взрослая при искусственном освещении) во вторую половину периода осеннего миграционного беспокойства проявился 4-суточный ритм локомоторной активности с максимумами 7–8, 12, 16, 19–20 и 24 октября (рис. 3).
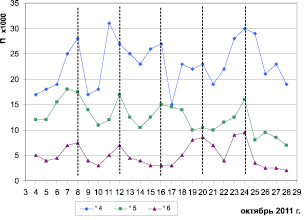
Рис. 3. Динамика уровня локомоторной активности трех самцов зябликов (№ 4 – sad, № 6 – ad при естественном освещение; № 5 – sad при искусственном освещении) в период с 4 по 28 октября 2011 г. По оси абсцисс – порядковый номер суток от начала исследования. По оси ординат – число прыжков за сутки
Рассуждая о возможных внешних синхронизаторах установленного 8-суточного ритма локомоторной активности для птиц, находившихся на естественном освещении, можно предположить чередование ясной и пасмурной погоды. Однако, судя по группе птиц, находившихся в условиях искусственного освещения, в синхронизации миграционной активности птиц участвует неизвестный внешний регулятор. В обоих случаях стимуляция беспокойства возможна только у подготовленных к миграции особей. Этим, вероятно, и объясняется наличие двух групп птиц, динамика локомоторной активности которых находится в противофазе, а акрофазы различаются между собой на четверо суток. Таким образом, каждые 4 суток возникает новая волна пролета мигрирующих птиц, наблюдаемая в природе.
Для многих видов птиц показано, что при миграции отмечается сезонное повышение уровня кортикостерона в сыворотке крови, так как этот гормон необходим для формирования физиологического статуса и мобилизации энергии в этот период [7, 12, 13]. Вероятнее всего, многодневное (инфрадианное) изменение уровня кортикостерона, подчиняющиеся влиянию внешних синхронизаторов, включено в регуляцию волнообразного изменения числа птиц, готовых к перелету.
Немногочисленные исследования многодневной динамики уровня глюкокортикоидных гормонов у млекопитающих указывают на существование около 4-суточного биоритма [5, 9, 11].
Ранее нами был выявлен 4-суточный инфрадианный ритм изменения уровня кортизола у человека и кортикостерона у лабораторных крыс, проявляющийся синхронно у разных особей и видов (с учетом формы суточной активности). Было установлено, что каждые 68–72 сут акрофаза инфрадианного ритма глюкокортикоидных гормонов у всех исследованных видов переходит на сутки вперед, обеспечивая кратность выявленного периода длине астрономического года [1]. Синхронность биоритмов позволила нам создать универсальный календарь наступления акрофаз и батифаз глюкокортикоидных гормонов. Установленная закономерность проявления биоритма у разных особей и видов указывает на наличие внешнего синхронизатора уровня глюкокортикоидных гормонов, природу которого пока трудно установить. Необходимо отметить, что в данном исследовании максимальный уровень локомоторной активности самцов зябликов наблюдался в дни, соответствующие расчетным датам акрофаз выявленного нами 4-суточного ритма изменения уровня глюкокортикоидных гормонов в сыворотке крови у крыс и человека.
При изучении колебаний уровня локомоторной активности у крыс-самцов Вистар нам удалось выявить 4-суточный период и положительную корреляцию с колебаниями уровня кортикостерона в сыворотке крови [2]. Несмотря на то, что птицы эволюционно разошлись с классом млекопитающих около 310 млн лет назад [9], можно предположить, что оба класса имеют сходные физиологические механизмы эндокринной регуляции локомоторной активности. В частности, в работе M.M. Landys и соавт. [10] выявлена положительная корреляция между уровнем кортикостерона и миграционной активностью, определяемой в неволе по уровню локомоторной активности. В качестве одного из претендентов на внешний синхронизатор I. Henshaw и соавт. [6] рассматривают геомагнитное поле, изменяющее гормональную секрецию кортикостерона, являющегося одним из наиболее важных гормональных факторов, обеспечивающих формирование и реализацию миграционного поведения.
Таким образом, в динамике уровня локомоторной активности самцов зябликов, содержавшихся в условиях неволи, во время осеннего миграционного состояния выявлен достоверный, регулярно проявляющийся инфрадианный ритм локомоторной активности с периодом 8 суток и наблюдающийся независимо от условий содержания – при естественном или искусственном освещении. У птиц, содержавшихся при естественном освещении, этот ритм синхронен у разных особей. У птиц, содержавшихся в условиях искусственного освещения, 8-суточный ритм локомоторной активности либо совпадал по фазе, либо находился в противофазе по отношению к фазе этого ритма в группе зябликов, находившихся при естественном освещении. У части птиц в конце миграционного периода установлена ритмичность изменения уровня локомоторной активности с периодом 4 суток. Выявленный характер динамики локомоторной активности птиц объясняет явление волнообразности пролета, наблюдаемое в природе при ежедневной регистрации числа летящих птиц.
Рецензенты:
Корабельникова Н.И., д.м.н., руководитель научно-организационного отдела, ФГБУ «Научно-исследовательский институт морфологии человека» РАМН, г. Москва;
Кондашевская М.В., д.б.н., руководитель группы функциональной морфологии стресса Научно-исследовательского института морфологии человека РАМН, г. Москва.
Работа поступила в редакцию 14.02.2013.
Библиографическая ссылка
Диатроптов М.Е. ИНФРАДИАННЫЙ РИТМ ЛОКОМОТОРНОЙ АКТИВНОСТИ У САМЦОВ ЗЯБЛИКОВ (Fringilla coelebs) В ПЕРИОД ОСЕННЕЙ МИГРАЦИИ // Фундаментальные исследования. 2013. № 4-5. С. 1139-1143;URL: https://fundamental-research.ru/en/article/view?id=31376 (дата обращения: 30.07.2026).