


Scientific journal
Fundamental research
ISSN 1812-7339
"Перечень" ВАК
ИФ РИНЦ = 1,798
ACTION OF POTASSIUM CHANNEL BLOCKER TETRAETHYLAMMONIUM IN SITUATION OF POTASSIUM DEPOLARIZATION IN MEMBRANE OF AMPHIBIAN MYELINATED NERVE FIBRES

Феномен калиевой деполяризации состоит в том, что в результате замены нормального раствора Рингера на гиперкалиевый происходит снижение мембранного потенциала, что сопровождается падением амплитуды потенциала действия (ПД). Блокирование калиевых каналов тетраэтиламмонием (ТЭА) устраняет вызванную избытком калия деполяризацию мембраны - и это несмотря на то, что при обычной концентрации ионов калия в наружном растворе ТЭА сам деполяризует мембрану [5, 6, 7].
В экспериментах, проведённых на миелинизированных нервных волокнах амфибий [10], было показано, что добавление ТЭА в наружный раствор, содержащий 30-60 мМ KCl, ведёт к восстановлению сниженного избытком ионов калия мембранного потенциала. Чуть позже [4] способность ТЭА восстанавливать сниженный гиперкалиевым раствором мембранный потенциал была подтверждена. Если сведения об уменьшении калиевой деполяризации под влиянием ТЭА в упомянутых работах [4, 10] были приведены, то относительно того, восстанавливалась ли под влиянием ТЭА генерация угнетённого гиперкалиевым раствором ПД, ничего не сказано. Этот пробел удалось устранить В.И. Беляеву и Б.И. Ходорову [1, 3], которые показали, что, помимо увеличения мембранного потенциала под влиянием ТЭА, в миелинизированных нервных волокнах амфибий происходит восстановление амплитуды ПД, до этого значительно уменьшенного гиперкалиевым раствором.
В экспериментах, проведённых на миелинизированных нервных волокнах амфибий [5], было установлено, что ТЭА приводил в нормальном растворе Рингера (концентрация ионов калия 2,5 мМ) к деполяризации мембраны вплоть до 8 мВ. В деполяризованных же с помощью гиперкалиевого (концентрация ионов калия от 10 до 117 мМ) раствора волокнах ТЭА смещал мембранный потенциал в противоположном направлении - вызывал гиперполяризацию мембраны приблизительно на 10 мВ.
В работе, выполненной на пучках миелинизированных нервных волокон амфибий Rana esculenta [6], установлено следующее. Если наружный раствор содержал от 0 до 2,5 мМ KCl, ТЭА приводил к деполяризации мембраны на 1-2,5 мВ, в то время как при концентрации ионов калия более 5 мМ ТЭА производил обратный эффект - происходила гиперполяризация (примерно на 10 мВ) мембраны. В работе, выполненной на одиночных миелинизированных нервных волокнах амфибий Rana esculenta и Xenopus laevis [7], показано, что ТЭА вызывает снижение мембранного потенциала в среднем на 5 мВ. В сенсорных волокнах степень деполяризации мембраны под влиянием ТЭА была меньше таковой моторных волокон. Добавление к раствору Рингера ТЭА вело к деполяризации мембраны, но если мембрану предварительно деполяризовали гиперкалиевым раствором, то ТЭА уменьшал эту деполяризацию в среднем на 7-15 мВ.
Различные нервные волокна по-разному реагируют на повышение концентрации ионов калия в среде [8, 9]. Разная чувствительность нервных волокон к калиевой деполяризации связывалась [8, 9] с тем, насколько хорошо были отпрепарированы волокна: в неповреждённых нервных волокнах увеличение концентрации ионов калия до 20 мМ приводило к небольшой и медленно нарастающей деполяризации, тогда как у плохо отпрепарированных волокон деполяризация возникала мгновенно, достигая уровня теоретического калиевого равновесного потенциала. По мнению автора [9], именно устойчивые к калиевой деполяризации волокна находятся в подобном in vivo состоянии.
В процессе изучения электрогенеза миелинизированных нервных волокон амфибий нами обнаружено два вида реакций волокон на избыток ионов калия в среде: в одних волокнах калиевая деполяризация развивается, а в других - нет. Представляется важным выявить, каким образом будет действовать блокатор калиевых каналов ТЭА в гиперкалиевой среде при ритмическом раздражении нервных волокон, реагирующих на избыток ионов калия деполяризацией мембраны, сведения о чём в литературе отсутствуют. Важность определения особенностей действия ТЭА в гиперкалиевой среде именно при ритмическом раздражении нервных волокон продиктована тем, что в процессе ритмической стимуляции можно проследить возможность суммирования следов от каждого одиночного нервного импульса - то есть смоделировать ситуацию, когда нервное волокно генерирует не один, а пачку нервных импульсов.
Материал и методы исследования
Эксперименты (n = 22) проводили на одиночных миелинизированных нервных волокнах седалищного нерва озёрной лягушки Rana ridibunda Pallas. Препаровку нервного волокна производили таким образом, что изолировали лишь интернодальную часть волокна, а перехват Ранвье, от которого в последующем отводили ПД, оставляли в нервном стволе. Подробное описание методики проведения экспериментов дано ранее [2]. О наступлении калиевой деполяризации в настоящей работе судили по уменьшению амплитуды ПД, а о её устранении - по увеличению амплитуды ПД. Судить о наступлении калиевой деполяризации по амплитуде ПД нам позволили данные В.И. Беляева, Б.И. Ходорова [1, 3], которые установили параллелизм между изменениями потенциала покоя и амплитуды ПД: в гиперкалиевом растворе под влиянием ТЭА происходило как увеличение амплитуды ПД, так и восстановление исходного потенциала покоя. Раствор Рингера, использованный в экспериментах, имел следующий состав (в мМ): NaCl - 111; KCl - 2,5; CaCl2 - 1,95; NaHCO3 - 1,2, HEPES («Sigma», США) - 10; рН 7,3. Эксперименты проводили при температуре 17-22 °С. Статистическая обработка экспериментальных данных осуществлялась с использованием критерия Стьюдента. Как и в предыдущей нашей работе, касающейся действия ТЭА в нормальном растворе Рингера [2], ТЭА («Sigma», США) использовался в концентрации 10 мМ. Гиперкалиевый раствор Рингера получали путём замены 30 мМ NaCl на такое же количество KCl.
Результаты исследования и их обсуждение
В ответ на приложение к нервному волокну одиночного деполяризующего стимула возникал ПД, сопровождающийся хорошо выраженной следовой деполяризацией (СД), амплитуда и длительность которой составили в среднем 2,12 ± 0,64 мВ и 175,27 ± 39,12 мс соответственно. Замена нормального раствора Рингера, содержащего 2,5 мМ К+, на гиперкалиевый (30 мМ К+) раствор вела к двум совершенно различным реакциям. В первом случае нервные волокна реагировали на повышение концентрации ионов калия в растворе падением амплитуды ПД и СД (35 % случаев), тогда как во втором случае амплитуда ПД достоверно не изменялась, а амплитуда СД существенно росла (65 % случаев). Итак, если в первом случае СД как по своей амплитуде, так и длительности уменьшалась (рис. 1, а), то во втором случае СД значительно увеличивалась (рис. 1, б). То, каким образом нервные волокна реагировали на избыток калия, зависело от исходного состояния волокна. В нормальном растворе Рингера амплитуда ПД нервных волокон, реагирующих на гиперкалиевый раствор по первому типу, составила 60,13 ± 16,64 мВ, тогда как у волокон, реагирующих по второму типу, она была равной 85,36 ± 11,87 мВ (различие статистически достоверно, р < 0,05).
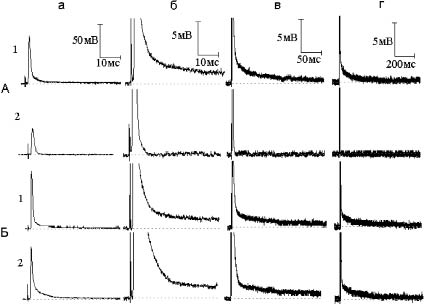
Рис. 1. Различные типы реакций миелинизированных нервных волокон на повышение концентрации ионов калия в наружном растворе с 2,5 до 30 мМ:
А - уменьшение следовой деполяризации (СД) под влиянием гиперкалиевого раствора.
1 - потенциал действия - ПД (а) и СД (б - г) в нормальном раство-ре Рингера. 2 - то же спустя три минуты после замены нормального раствора Рингера на гиперкалиевый. Б - увеличение СД под действием гиперкалиевого раствора. 1 - ПД (а) и СД (б - г) в нормальном растворе Рингера. 2 - то же спустя четыре минуты после замены нормального раствора Рингера на гиперкалиевый
Калиевая деполяризация изучалась на тех волокнах, которые отвечали на избыток ионов калия в среде подавлением ПД. Эксперимент, в котором замена нормального раствора Рингера на гиперкалиевый привела к калиевой деполяризации, представлен на рис. 2 и 3. Рис. 2 иллюстрирует изменение амплитудно-временных характеристик ПД и СД при одиночном раздражении нервного волокна: избыток ионов калия привёл к уменьшению амплитуды ПД и СД, добавление же к гиперкалиевому раствору ТЭА вызвало восстановление ПД и значительное увеличение амплитуды и длительности СД. Увеличение амплитуды и длительности СД под влиянием ТЭА можно также наблюдать и в нормальном (2,5 мМ К+) растворе Рингера [2].
На рис. 3 можно проследить изменения следовых потенциалов в этом эксперименте при ритмическом раздражении нервного волокна. Падение амплитуды ПД и уменьшение амплитуды и длительности СД, вызванное гиперкалиевым раствором, привело к тому, что суммация СД в ритмическом ряду была существенно уменьшена, а посттетаническая деполяризация (ПТД), представляющая собой замедленное возвращение мембранного потенциала к исходному уровню после прекращения ритмической стимуляции, практически исчезла. Добавление к гиперкалиевому раствору ТЭА вызвало увеличение как амплитуды деполяризационного плато, так и амплитуды и длительности ПТД.
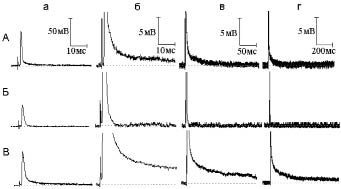
Рис. 2. Влияние, оказываемое тетраэтиламмонием на электрическую активность нервного волокна при одиночном раздражении в гиперкалиевом (30 мМ) растворе Рингера:
А - потенциал действия (а) и следовая деполяризация (б - г) в нормальном растворе Рингера.
Б - то же через шесть минут действия гиперкалиевого раствора. В - то же через три минуты после добавления к гиперкалиевому раствору тетраэтиламмония
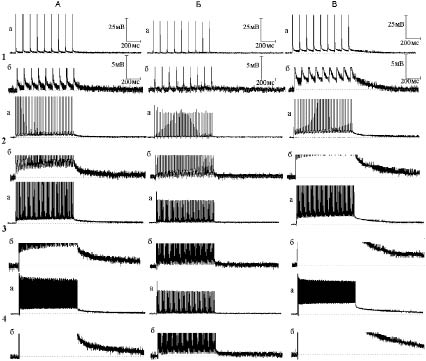
Рис. 3. Эффект совместного действия тетраэтиламмония (ТЭА) и гиперкалиевого (30 мМ) раствора Рингера на следовые потенциалы при ритмическом раздражении нервных волокон
(тот же опыт, что и на рис. 2):
А - следовые потенциалы при ритмической стимуляции нервного волокна частотой 10 (1), 50 (2), 100 (3) и 300 (4) Гц в нормальном растворе Рингера. Б - то же через шесть минут действия гиперкалиевого раствора. В - то же через три минуты после добавления к гиперкалиевому раствору ТЭА
Можно было бы подумать, что такое колоссальное увеличение длительности ПТД вызвано сугубо одним действием ТЭА, но это не так. Столь колоссального увеличения длительности ПТД добиться в экспериментах, в которых ТЭА действует в нормальном растворе Рингера, невозможно. При частоте раздражения 50 Гц длительность ПТД в растворе Рингера, содержащем 2,5 мМ К+ и 10 мМ ТЭА, составила 0,45 ± 0,12 мс, в то время как в растворе, содержащем 30 мМ К+ и 10 мМ ТЭА, в серии экспериментов с нервными волокнами, чувствительными к гиперкалиевому раствору, она составила 1,17 ± 0,41 мс (различия статистически достоверны, p < 0,05, n = 7). Чтобы изменения были столь велики, ТЭА должен действовать в гиперкалиевом, а не в нормальном растворе Рингера. ПТД, регистрируемая при действии ТЭА в гиперкалиевой среде, напоминает ПТД, получаемую в гиперкалиевом растворе без ТЭА - и именно у тех нервных волокон, которые отвечают на избыток ионов калия в среде увеличением СД. Длительность ПТД в гиперкалиевом растворе в волокнах, не чувствительных к избытку ионов калия, при частоте раздражения 50 Гц составила 1,60 ± 0,34 мс. На рис. 4 и 5 приведены следовые потенциалы, получаемые у такого типа волокон при одиночном и ритмическом раздражении соответственно. Как можно видеть, характер изменения ПТД под влиянием избытка ионов калия (см. рис. 5) сходен с таковым в гиперкалиевой среде, содержащей ТЭА (рис. 3).
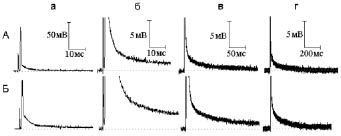
Рис. 4. Влияние, оказываемое гиперкалиевым (30 мМ) раствором на электрическую активность миелинизированного нервного волокна при одиночном раздражении:
А - потенциал действия (а) и следовая деполяризация (б - г) в нормальном растворе Рингера.
Б - то же через семь минут действия гиперкалиевого раствора
Тот факт, что в настоящей работе выявлено два типа реакций миелинизированных нервных волокон на повышение концентрации ионов калия, позволяет оценивать физиологическое состояние нервного проводника - калиевая деполяризация возникала в тех волокнах, амплитуда ПД которых колебалась в пределах от 30 до 80 мВ (60,13 ± 16,64 мВ). Устойчивые к гиперкалиевому раствору волокна характеризовались амплитудой ПД в интервале от 71 до 106 мВ (85,36 ± 11,87 мВ). Важно, что совершенно противоположная реакция волокон первого и второго типа указывает на то, что у неповреждённой мембраны имеется возможность сохранять потенциал покоя неизменным и предотвращать наступление калиевой деполяризации. Причину роста СД в гиперкалиевом растворе у неповреждённых нервных волокон мы видим в уменьшении концентрационного градиента для ионов калия на фоне отсутствия изменений потенциала покоя (либо сравнительно небольшого его изменения) мембраны волокна при повышении наружной концентрации ионов калия. В противоположность этому уменьшение потенциала покоя нервной мембраны должно вести как к падению амплитуды ПД, так и уменьшению величины СД.
Различные реакции нервных волокон на повышение концентрации ионов калия в среде согласуются с ранее известными исследованиями [8, 9], в которых развитие калиевой деполяризации зависело от состояния нервного волокна. В тех волокнах, которые находились в хорошем физиологическом состоянии, калиевая деполяризация развивалась медленно, тогда как в волокнах, находящихся в плохом состоянии, калиевая деполяризация развивалась стремительно.
Эксперименты, представленные в настоящей работе, позволили выявить особенности действия ТЭА в гиперкалиевом растворе. Гиперкалиевый раствор в отсутствие ТЭА приводит к двум совершенно различным реакциям. В волокнах, чувствительных к избытку ионов калия, гиперкалиевый раствор уменьшает величину СД, деполяризационного плато и ПТД. В нечувствительных к избытку ионов калия волокнах (у которых под влиянием гиперкалиевого раствора потенциал покоя либо совсем не изменяется, либо его изменения незначительны), СД, деполяризационное плато и ПТД в гиперкалиевом растворе значительно возрастают. Устраняя калиевую деполяризацию в чувствительных к избытку ионов калия волокнах (то есть восстанавливая исходный мембранный потенциал), ТЭА даёт возможность наблюдать увеличение следовых потенциалов, по характеру сходное с увеличением следовых потенциалов, наблюдаемых у нечувствительных к гиперкалиевому раствору волокон. Заметим, что ТЭА увеличивает следовые потенциалы посредством блокирования калиевых каналов, а гиперкалиевый раствор - посредством уменьшения концентрационного градиента для ионов калия. И гиперкалиевый раствор, и ТЭА в данном случае действуют однонаправлено - вызывают увеличение следовых потенциалов, причём при их совместном действии возникает значительная трудность в разделении производимых ими эффектов. Мы столкнулись с тем, что в чувствительных к избытку ионов калия волокнах под влиянием ТЭА происходит значительное увеличение длительности ПТД, и это увеличение можно было бы ошибочно приписать только одному действию ТЭА. Имея в распоряжении информацию, что ТЭА в нормальном растворе Рингера производит не столь значительное увеличение ПТД, нами намеренно поставлены эксперименты в гиперкалиевой среде без ТЭА в волокнах, не чувствительных к калиевой деполяризации. Именно эти эксперименты позволили отделить увеличение ПТД, производимое ТЭА, от такового, производимого гиперкалиевым раствором. Стало ясно, что столь значительное увеличение ПТД под влиянием ТЭА можно получить лишь в гиперкалиевой среде.
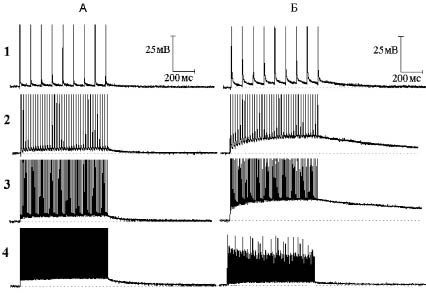
Рис. 5. Влияние гиперкалиевого (30 мМ) раствора на следовые потенциалы при ритмическом раздражении нервного волокна (тот же опыт, что на рис. 4):
А - следовые потенциалы при ритмической стимуляции нервного волокна частотой 10 (1), 50 (2), 100 (3) и 300 (4) Гц в нормальном растворе Рингера. Б - то же через семь минут после замены нормального раствора Рингера на гиперкалиевый
Блокаторы калиевых каналов, предотвращая движение ионов калия через мембрану, способствуют восстановлению сниженного гиперкалиевым раствором мембранного потенциала. Поскольку проводящих каналов под влиянием блокаторов становится меньше, значит меньшее количество каналов будет участвовать в установлении новой разности потенциалов на мембране при повышении концентрации ионов калия - то есть снижение мембранного потенциала будет замедляться.
Выводы
1. Миелинизированные нервные волокна амфибий отвечают на замену нормального раствора Рингера (2,5 мМ К+) на гиперкалиевый (30 мМ К+) двумя совершенно противоположными реакциями. В первом случае происходит падение амплитуды ПД и сопровождающее это уменьшение амплитуды и длительности СД, во втором же случае амплитуда ПД достоверно не изменяется, а амплитуда и длительность СД растут.
2. В волокнах, чувствительных к избытку ионов калия, гиперкалиевый раствор уменьшает величину СД, деполяризационного плато и ПТД. В нечувствительных к избытку ионов калия волокнах (у которых под влиянием гиперкалиевого раствора потенциал покоя либо совсем не изменяется, либо его изменения незначительны), СД, деполяризационное плато и ПТД в гиперкалиевом растворе значительно возрастают.
3. В тех волокнах, в которых в гиперкалиевом растворе наблюдается калиевая деполяризация (снижение амплитуды ПД), введение в гиперкалиевый раствор ТЭА (10 мМ) ведёт к восстановлению сниженного избытком ионов калия ПД, а также увеличению амплитуды и длительности СД.
4. Добавление к гиперкалиевому раствору ТЭА вызывает значительное увеличение длительности ПТД, получаемой при ритмическом раздражении нервных волокон, чувствительных к избытку ионов калия. Чтобы изменения были столь велики, ТЭА должен действовать в гиперкалиевом, а не в нормальном растворе Рингера. Устраняя калиевую деполяризацию в чувствительных к избытку ионов калия волокнах, ТЭА даёт возможность наблюдать увеличение следовых потенциалов, характерное для нечувствительных к гиперкалиевому раствору волокон, проявляющих увеличение следовых потенциалов в гиперкалиевом растворе в отсутствие ТЭА.
Список литературы
- Беляев В.И. Сопоставление изменений электрической активности одиночного перехвата Ранвье при повышении концентрации ионов калия в среде и действии новокаина // Бюллетень экспериментальной биологии и медицины. - 1964. - № 12. - С. 13-17.
- Изменение электрогенеза миелинизированных нервных волокон амфибий под действием тетраэтиламмония / И.В. Кузнецова, Д.А. Евстигнеев, Н.В. Глухова, В.П. Глухов //
Современные наукоёмкие технологии. - 2008. - № 2. - С. 22-29. - Ходоров Б.И., Беляев В.И. Физиологический электротон одиночного перехвата Ранвье в условиях воздействия ионов тетраэтиламмония // Биофизика клетки. - М.: Наука, 1965. - С. 159-174.
- Lüttgau H.C. Das Kalium-Transportsystem am Ranvier-Knoten isolierter markhaltiger Nervenfasern // Pflügers Archiv. - 1960. - Vol. 271. - P. 613-633.
- Schmidt H. Zur Wirkung von Tetraäthylammoniumchlorid (TEA) auf das Membranpotential markhaltiger Nervenfasern // Pflügers Archiv. - 1963. - Vol. 278. - P. R4-R5.
- Schmidt H. Die Wirkung von Tetraäthylammoniumchlorid auf das Membranpotential und den Membranwiderstand von Bündeln markhaltiger Nervenfasern // Pflügers Archiv. - 1965. - Vol. 282. - P. 351-361.
- Schmidt H., Stämpfli R. Die Wirkung von Tetraäthylammoniumchlorid auf den einzelnen Ranvierschen Schnürring // Pflügers Archiv. - 1966. - Vol. 287. - P. 311-325.
- Stämpfli R. Die Strom-Spannungs-Charakteristik der erregbaren Membran eines einzelnen Schnürrings und ihre Abhängigkeit von der Ionenkonzentration // Helv. Physiol. et Pharmacol. Acta. - 1958. - Vol. 16. -P. 127-145.
- Stämpfli R. Is the resting potential of Ranvier nodes a potassium potential? // Annals New York Academy of Sciences. - 1959. - Vol. 81. - P. 265-284.
- Tasaki I. Demonstration of two stable states of the nerve membrane in potassium-rich media // J. Physiol. - 1959. - Vol. 148. - P. 306-331.
Рецензенты:
Песков А.Б., д.м.н., профессор, декан факультета постдипломного, дополнительного и высшего сестринского образования института медицины, экологии и физической культуры Ульяновского государственного университета, г. Ульяновск;
Русанов А.М., д.б.н., профессор, декан химико-биологического факультета Оренбургского государственного университета, г. Оренбург.
Библиографическая ссылка
Кузнецова И.В, Евстигнеев Д.А, Глухова Н.В ДЕЙСТВИЕ БЛОКАТОРА КАЛИЕВЫХ КАНАЛОВ ТЕТРАЭТИЛАММОНИЯ В УСЛОВИЯХ КАЛИЕВОЙ ДЕПОЛЯРИЗАЦИИ МЕМБРАНЫ МИЕЛИНИЗИРОВАННЫХ НЕРВНЫХ ВОЛОКОН АМФИБИЙ // Фундаментальные исследования. 2011. № 7. С. 196-202;URL: https://fundamental-research.ru/en/article/view?id=26757 (дата обращения: 01.07.2026).