



Отдаленная гибридизация между родами тритикале и пшеницы дает перспективный материал как для селекции, так и для изучения процессов, которые происходят при формообразовании [4]. При таких скрещиваниях происходят рекомбинации как между геномами пшеницы и ржи, так и между геномами пшениц, принадлежащих разным видам рода Triticum. Самым перспективным и широко используемым типом таких скрещиваний являются скрещивания гексаплоидных тритикале с мягкой пшеницей. Но для изучения процессов формообразования интересны также гибриды гексаплоидных тритикале с тетраплоидной пшеницей, родительские виды которых различаются как по геномному составу пшеничного компонента, так и отсутствием буферной роли D-генома [10, 9]. Работ, посвященных этому типу скрещиваний, относительно немного и в них использованы тетраплоидные пшеницы T. durum и T. turgidum [6, 8, 10].
Целью нашей работы явилось создание исходного материала и изучение формообразовательных процессов в гибридных популяциях, полученных при скрещивании пшенично-ржаного амфиплоида ABDR с тетраплоидными пшеницами T. dicoccum и T. paleocolchicum.
Материалы и методы исследования
Объектом исследования являлись тритикально-шеничные гибриды F1-F5, которые изучали с 2009 по 2013 год. В гибридизации был использован пшенично-ржаной амфиплоид ABDR (2n = 6х = 42, геном AАBВD/R), выделенный из гибридной популяции синтетической пшеницы ABD (T. durum×Ae. squarrosa) c сорно полевой рожью Secale cereale ssp. segetale (2n = 2х = 14, геном RR) [1]. Пшеница была представлена двумя видами: T. dicoccum var. rufum (2n = 4х = 28, геном AABB) и T. paleocolchicum Men. (2n = 4х = 28, геном AABB). Гибридизация ABDR с тетраплоидными пшеницами была проведена по схеме реципрокного скрещивания: тритикале/пшеница (прямые) и пшеница/тритикале (обратные).
Кастрацию колосьев проводили по общепринятой методике. Определяли завязываемость и жизнеспособность гибридных зёрен, и генетическую совместимость ABDR и тетраплоидных пшениц
( ).
).
Посев образцов и их гибридов проводили вручную, растения убирали с корнями. Растения гибридных популяций F1-F5 анализировали по морфологическим признакам и элементам продуктивности в сравнении с исходными родительскими формами. Показатель степени фенотипического доминирования признаков растений в первом поколении гибридов определяли по формуле G.M. Beil и R.E. Atkins (1965) [7]. Начиная с третьего поколения, у гибридных растений по отношению к пшеничному родителю определяли степень и частоту положительных и отрицательных трансгрессий изучаемых признаков по методике Воскресенской‒Шпота (1967) [2].
Статистическую обработку данных проводили в соответствии со стандартными методиками [3], с использованием компьютерных программных обеспечений Microsoft Excel 2010 и IBM SPSS Statistics (Version 20).
Результаты исследования и их обсуждение
Результаты скрещивания пшенично-ржаного амфиплоида ABDR с тетраплоидными пшеницами по 2 комбинациям приведены в табл. 1. Как видно из таблицы, комбинационная совместимость амфиплоида ABDR с тетраплоидной пшеницей различалась незначительно. Большая совместимость была в комбинации ABDR×T. dicoccum var. rufum (17,32 %), несколько меньше в комбинации ABDR×T. paleocolchicum Men. (14,14 %). Исходя из литературных данных [5], успех гибридизации в скрещиваниях тритикале с тетраплоидной пшеницей зависит как от климатических условий, так и от направления скрещивания, приводящих к достоверным различиям в образовании и жизнеспособности гибридных зерновок при прямых и обратных скрещиваниях. В отличие от результатов венгерского ученного Киша [5], в климатических условиях Апшеронской опытной станции, где проводились наши опыты, гибридные семена завязывались лучше при скрещивании ABDR с гексаплоидной пшеницей, чем с твёрдой, однако всхожесть в случае с последними была также ниже. Так, в нашем материале в среднем завязываемость семян в комбинации ABDR×4х пшеница составила 9,98 %, а в обратных скрещиваниях в два раза больше, чем в прямых, и составила в среднем 20,13 %. Эти показатели почти в два раза меньше показателей завязываемости семян при скрещивании ABDR с гексаплоидной пшеницей, где в среднем при прямых скрещиваниях она составила 16,69 %, а при обратных – 36,39 % соответственно.
Таблица 1
Скрещиваемость пшенично-ржаного амфиплоида ABDR с тетраплоидной пшеницей
|
№ п/п |
Комбинация скрещивания |
Опылено цветков |
Завязалось гибридных зерен |
Всхожесть, % |
Полевая выживаемость, % |
Совместимость, % |
|
|
число |
% |
||||||
|
1. |
ABDR×T. dicoccum |
102 |
12 |
11,76 |
25 |
33,33 |
17,32 |
|
2. |
T. dicoccum×ABDR |
164 |
36 |
21,95 |
0 |
– |
0 |
|
3. |
ABDR×T. paleocolchicum |
122 |
13 |
10,66 |
20 |
33,33 |
14,14 |
|
4. |
T. paleocolchicum×ABDR |
153 |
28 |
18,30 |
0 |
– |
0 |
При этом жизнеспособность зерновок проявляет такую же закономерность, как и при скрещиваниях ABDR с гексаплоидными пшеницами, а именно в нашем материале в комбинациях 4х пшеница×ABDR не получено гибридных растений, а в комбинациях ABDR×4х пшеница она составила в среднем 22.5 %. Таким образом, имелись резкие реципрокные различия при скрещивании ABDR с тетраплоидной пшеницей. Это согласуется с литературными данными о том, что если при гибридизации с мягкой пшеницей в обратной комбинации все же образуются немногие жизнеспособные семена, то при участии твердой пшеницы отмечается полная постгамная несовместимость. Гибридные зерновки обратной комбинации не всходили даже при проращивании на питательной агаровой среде [5].
Гибриды F1 в обоих комбинациях морфологически были промежуточными, с тонкими прочными стебляим и узкими листьями, имели низкую кустистость. Колосья были неопушенные, с черными остьями и со слабым опушением под колосом. В комбинации ABDR×T. dicoccum (рис. 1) колосья имели тенденцию к ветвистости, что отмечалось и другими ученными в комбинациях скрещивания гексаплоидного тритикале с T. durum и T. turgidum [10, 6]. Характеристика наследования количественных признаков у гибридов ABDR с тетраплоидными пшеницами в ряду поколений дана в таблице 2. Отмечено, что по высоте растений (ВР) у гибридов F1 наблюдалось неполное доминирование высокорослости (0,54 ≤ hp ≤ 0,69). По признаку «длина колоса» (ДК) гибриды F1 превосходили пшеничных родителей, наблюдалось частичное доминирование (–0,33 ≥ hp ≥ –0,41) малой длины колоса. По числу колосков в колосе у гибридов F1 в комбинации ABDR×T. dicoccum наблюдалось частичное доминирование (0 > hp ≥ –0,22), а в комбинации ABDR×T. paleocolchicum сверхдоминирование (hp = –8) малого числа колосков в колосе. По фертильности колоса (ФК) гибриды F1 уступали исходным формам, в среднем по комбинациям она равнялась 3,3 и 4,3 %. При этом всхожесть семян, полученных от растений F1, в обеих комбинациях была очень низкой. Так, для комбинации ABDR×T. paleocolchicum она составила 15.38 %, а для комбинации ABDR×T. dicoccum –12 %. При этом полевая выживаемость гибридов F2 от скрещивания ABDR с тетраплоидными пшеницами оказалась также низкой. В полевых условиях выжило только одно растение из комбинации ABDR×T. dicoccum, остальные растения из этой комбинации погибли в фазе трёх листьев. Два гибридных растения F2 из комбинации ABDR×T. paleocolchicum достигли конца вегетации, но оказались полностью стерильными. Выжившее растение F2 в комбинации ABDR×T. dicoccum характеризовалась высокой продуктивной кустистостью (21 стеблей, из которых 19 имели более или менее озерненные колосья) и недетерминированным летним кущением, которое продолжалось вплоть до уборки. Колосья этого растения были пшеничного типа, менее ломкие по сравнению с пшеничным родителем, красного цвета, неопушенные, с черными остьями, без опушения под колосом. Высота этого растения, как и стерильных растений из комбинации ABDR×T. paleocolchicum, оказалось ниже этого показателя у родительских растений, как исходных, так и растений F1, т.е. наблюдалась тенденция к низкорослости. Колосья стерильных растений комбинации ABDR×T. paleocolchicum были тритикального типа, неопушенные, остистые, с опушением под колосом. Как видно из табл. 2, ФК гибридного растения F2 из комбинации ABDR x T. dicoccum оказалось достаточно высокой, было получено 284 зерна, из которых проросли 218 (в лаборатории). Полевая выживаемость этих растений оказалась низкой и составила 55 %, отчасти из-за неблагоприятных погодных условий, сложившихся как в первой половине вегетации (несвойственная высокая температура для осенних месяцев 2010 г. и для первых месяцев зимы 2011 г.), так и во второй половине развития растений (высокий естественный инфекционный фон жёлтой ржавчины и мучнистой росы). Все гибридные растения третьего поколения фенотипически были подразделены на растения пшеничного, тритикального и промежуточного типа, которые составили соответственно 73, 23 и 4 % от общего числа популяции. Начиная с F3 в незначительном количестве отмечены формы, сходные с другими видами пшениц: T. polonicum и T. turgidum (рис. 2). Высота гибридных растений F3 в среднем для всей популяции составила 83 см, для фенотипической группы пшеничного типа 86 см, а для тритикального и промежуточного типа – 72 и 71 см соответственно. При этом самые высокорослые растения (более 123 см) вошли только в фенотипическую группу пшеничного типа, а низкорослые – до 123 и 84 см ‒ в группу промежуточного и тритикального типа. Степень трансгрессии по ВР у гибридов F3 колебалась от –56 до 46 %, а частота трансгрессии составила 80 %. Длина колоса у всех перечисленных фенотипических групп в среднем была равна 12 см, но количество колосков у тритикального и промежуточного типа оказалось больше на 2–3 шт. по сравнению с пшеничным морфотипом. Положительной трансгрессии по признаку «длина колоса» не наблюдалось, отрицательная трансгрессия колебалась от –10 до 49 %, а частота трансгрессии составила 50 %. Продуктивность растений по фенотипическим группам у гибридов F3 оказалось различной: от 0 до 99 % для пшеничной группы, от 0 до 48 % для тритикальной и от 0 до 12 % ‒ для промежуточной группы. Выявлена положительная корреляционная связь между ВР и ФК (r = 0,399, P < 0,01), а также между ДК и ЧК (r = 0,415, P < 0,01). Фенотипическое разделение оказалось следующим: 92 % для пшеничного типа, 2 % ‒ для тритикального и 4 % ‒ для промежуточного типа. Высота гибридных растений F4 в среднем для всей популяции по сравнению с F3 возросла на 20 см и составила 102 см, а для фенотипических групп – 103 см для пшеничного типа, 87 см ‒ для тритикального и 98 см ‒ для промежуточного типа. Степень трансгрессии по ВР у гибридов F4 колебалась от –65 до 13 %, а частота трансгрессии составила 40 %. Длина колоса в среднем для всей популяции F4, как и для F3, составила около 12 см, с самым широким варьированием у растений пшеничного морфотипа – от 4,5 до 23,5 см. В отличие от гибридов F3 в F4 наблюдалась положительная трансгрессия от 5до 12 % по признаку «длина колоса» с частотой трансгрессии 1 %. Положительной трансгресии по ЧК не наблюдалось. Продуктивность растений по фенотипическим группам у гибридов F4 оказалась различной: от 1 до 100 % ‒ для пшеничной группы, от 15 до 57 % ‒ для тритикальной и от 3 до 97 % для промежуточной группы. Выявлена положительная корреляционная связь между ВР и ФК (r = 0,114, P < 0,05), между ВР и ДК (r = 0,159, P < 0,01), а также между ДК и ЧК (r = 0,131, P < 0,01), а отрицательная корреляционная связь между ДК и ФК (r = –0,105, P < 0,05). Фенотипическое подразделение на группы растений F5 оказалось следующим: 97 % ‒ для пшеничного типа, 1 % ‒ для тритикального и 2 % для промежуточного типа. Высота гибридных растений F5 в среднем для всей популяции по сравнению с F4 возросла на 10 см и составила 112 см, а для фенотипических групп – 115 см для пшеничного типа, 68 см ‒ для тритикального и 105 см ‒ для промежуточного типа. Степень трансгрессии и частота трансгрессии по ВР у гибридов F5 оказалась примерно такой же, как у F4, что свидетельствует об относительной стабилизации популяции. Длина колоса в среднем для всей популяции F5 составила 13 см, с самым широким варьированием у растений пшеничного морфотипа – от 4 до 30 см. Наблюдалась положительная трансгрессия от 2,4 до 40 % по признаку «длина колоса» с частотой трансгрессии 8 %. Но положительной трансгресии по ЧК в F5, как и F4, не наблюдалось. Продуктивность растений по фенотипическим группам у гибридов F5 оказалась различной: от 4 до 100 % (в среднем 61 %) для пшеничной группы, от 20 до 63 % (в среднем 38 %) ‒ для тритикальной и от 4 до 100 % ‒ для промежуточной группы. Выявлена положительная корреляционная связь между ВР и ФК (r = 0,328, P < 0,01), между ДК и ЧК (r = 0,273, P < 0,01), а также между ВР и ЧК (r = 0,135, P < 0,05).

Рис. 1. Гибриды F1 комбинации ABDR×T. dicoccum
Из гибридов F4-F5 выделен ряд трансгрессивных низкорослых (41–70 см) и высокорослых (95–103 см) форм твёрдой пшеницы, отличающихся хорошо озернённым колосом и выполненным зерном. Следует отметить, что по сравнению с поздними поколениями комбинаций ABDR×6х пшеница, в поздних комбинациях ABDR×T. dicoccum промежуточная и тритикальная фенотипические группы были малочисленными. Из последних фенотипических групп по некоторым биологическим свойствам отобраны 4 низкорослые (55–86 см) формы для дальнейшего изучения.
Таким образом, было установлено, что при скрещивании ABDR с тетраплоидной пшеницей имелись резкие реципрокные различия. Гибриды F1 были низкорослы и длинноколосы по сравнению с пшеничным родителем. В последующих генерациях в результате расщепления по морфологическим признакам и биологическим свойствам появляются трансгрессивные низкорослые и высокорослые формы твёрдой пшеницы и тритикале. Также выявлено, что формообразовательный процесс у гибридов амфиплоида ABDR с тетраплоидной пшеницей во многих поколениях протекает с увеличением доли растений пшеничного типа из-за быстрой элиминации ржаных хромосом, в результате чего большая часть растений гибридной популяции представляет собой сложную мозаику межвидовых гибридов пшеницы.
Таблица 2
Сравнительная характеристика наследования количественных признаков в ряду поколений (F1-F5) у гибридов ABDR с тетраплоидной пшеницей
|
Поколение/Родители |
Гибридные омбинации/Родительские формы |
ВР, см
|
ДК, см
|
ЧК, шт.
|
ПК,
|
ФК, %
|
|
P |
ABDR |
100 |
21 |
41 |
1,90 |
93 |
|
T. dicoccum var. rufum |
139 |
11,8 |
23 |
1,86 |
100 |
|
|
T. paleocolchicum Men. |
132 |
11 |
40 |
3,55 |
100 |
|
|
F1 |
ABDR×T. dicoccum |
130 |
14,5 |
30 |
2 |
3,3 |
|
ABDR×T. paleocolchicum |
127 |
15,0 |
33 |
2,13 |
4,3 |
|
|
F2 |
ABDR×T. dicoccum |
112 |
13,4 |
26 |
1,86 |
76,8 |
|
ABDR×T. paleocolchicum |
67 |
10 |
24 |
2,3 |
0 |
|
|
F3 |
ABDR×T. dicoccum |
82,49 ± 2,25 (44–173)* |
11,68 ± 0,29 (6–19) |
22,08 ± 0,39 (13–32) |
1,96 ± 0,05 (1,19–3,43) |
10,41 ± 2,18 (0-99) |
|
F4 |
ABDR×T. dicoccum |
101,9 ± 1,13 (35–173) |
11,53 ± 0,17 (4–23,50) |
21,56 ± 0,16 (9–32) |
2,02 ± 0,03 (0,89–4,26) |
53,71 ± 1,47 (1–100) |
|
F5 |
ABDR×T. dicoccum |
112,08 ± 1,99 (35–173) |
13,23 ± 0,33 (3,80–30) |
25,83 ± 0,24 (15–38) |
2,17 ± 0,05 (0,83–4,80) |
58,46 ± 1,78 (3,70–100) |
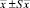
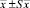
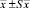
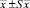
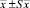
Примечание. *В скобках – границы варьирования.




Рис. 2. Гибридные формы в F4 и F5, сходные с другими видами пшениц: T. polonicum и T. turgidum
Рецензенты:
Шыхлински Г.М., д.б.н., старший научный сотрудник, заведующий отделом охраны растений, Институт генетических ресурсов НАНА, г. Баку;
Алиев Р.Т., д.б.н., профессор, заведующий отделом физиологии растений, Институт генетических ресурсов НАНА, г. Баку.
Работа поступила в редакцию 25.12.2013.
Библиографическая ссылка
Мехтиева С.П., Аминов Н.Х. ФОРМООБРАЗОВАТЕЛЬНЫЙ ПРОЦЕСС ПРИ СКРЕЩИВАНИЯХ ГЕКСАПЛОИДНОГО ТРИТИКАЛЕ С ПОЛБОЙ // Фундаментальные исследования. 2013. № 11-6. С. 1191-1196;URL: https://fundamental-research.ru/ru/article/view?id=33275 (дата обращения: 02.08.2026).