


Scientific journal
Fundamental research
ISSN 1812-7339
"Перечень" ВАК
ИФ РИНЦ = 1,798
THE STUDY OF THE INFLUENCE OF THE SUM OF TRITERPENE ACIDS FROM THE FRUITS OF SEA BUCKTHORN (HIPPOPHAë RHAMNOIDES L.) AND CRANBERRY (VACCINIUM OXYCOCCOS L.) ON LIPOLYSIS IN ADIPOSE TISSUE OF LABORATORY ANIMALS AT COURSE INTRODUCTION

Формирование первичных и вторичных гиперлипидемий сопровождается изменениями показателей, характеризующих состояние липидного обмена, в основе которых лежат различные механизмы – стимуляция выхода липидов из депо в кровь, нарушение экскрекции и утилизации холестерина печенью, снижение активности постгепариновой липопротеинлипазы крови и другие [2].
Стимуляция липолиза и выброс продуктов гидролиза тканевых липидов зависит от состояния механизмов регуляции активности липолитических гидролаз. Различают несколько типов липаз, активность которых регулируется различными факторами: липаза, активируемая адреналином; липаза, действующая в нестимулированной ткани; липаза липопротеидная, активность которой повышается при инкубации с гепарином. Жиромобилизующая липаза осуществляет гидролиз триглицеридов, обеспечивает поступление в кровь неэтерефицированных жирных кислот с последующим их использованием в качестве энергетического материала. Липазу жировой ткани кроме адреналина активируют норадреналин, соматотропный гормон, адренокортикотропный гормон. Липопротеидная липаза обладает как липолитическим, так и липосинтетическим действием в отношении триглицеридов жировой ткани. Кроме гепарина активность липопротеидлипазы повышает добавление в инкубационную среду инсулина и глюкозы. У человека с нормальным обменом веществ, не страдающего ожирением, и та и другая липаза, будучи достаточно активными, в известной мере уравновешивают процессы липогенеза и липосинтеза. На первой стадии внутриклеточного липолиза происходит гидролиз триглицеридов до диглицеридов, и эта медленная стадия лимитирует скорость всего процесса. Стимулирует его цАМФ, образующийся аденилатциклазой в ответ на гормональную стимуляцию. Мембрана жировой клетки обладает двумя типами рецепторов гормонов. С первыми взаимодействуют катехоламины (адреналин или норадреналин), усиливающие образование цАМФ, а значит и липолиз, со вторыми взаимодействует инсулин, который снижает концентрацию цАМФ, противодействуя активации аденилатциклазы стимулирующими гормонами, и угнетает таким образом липолиз.
Соединения, обладающие гиполипидемическим действием, способны восстанавливать нарушенные механизмы регуляции липидного обмена [1]. Среди растительных веществ с обнаруженной гиполипидемической активностью особенно интересным представляется исследование возможного влияния на активность тканевых липолитических ферментов соединений близких по строению к некоторым эндогенным гуморальным регуляторам. Так, исследуемые нами объекты предположительно могут реализовывать свой гиполипидемический эффект благодаря гормоноподобному действию, обусловленному родством структуры урсоловой и олеаноловой кислот со стероидными регуляторами, активно участвующими в регуляции липидного обмена (рисунок).
Целью данной работы явилось изучение влияния курсового введения тритерпеноидов облепихи (ТО) и тритерпеноидов клюквы (ТК) на интенсивность катехолстимулированного липолиза в жировой ткани.
Материалы и методы исследования
Сумма тритерпеновых кислот облепихи (ТО) и сумма тритерпеновых кислот клюквы (ТК) (предоставлены ООО «СиНаМ») – аморфный порошок от белого до желто-кремового цвета. Содержание суммы урсоловой и олеаноловой кислот не менее 75 %.
Исследования выполнены на 32 крысах-самцах линии Wistar массой 230–250 г. Источник получения животных – ФГУП «Питомник лабораторных животных «Рапполово» РАМН. Животные содержались на стандартном пищевом рационе со свободным доступом к корму (Полнорационный комбикорм рецепт ПК-120 для содержания лабораторных животных, ГОСТ Р 50258-92, производитель ООО «Лабораторкорм») и воде. После 14-дневного карантина животные распределялись по группам по 8 животных в каждой. ТО и ТК вводили перорально раз в сутки в виде водной суспензии (стабилизатор твин-80) в дозе 100 мг/кг в течение 30 суток. Контрольной группе животных вводили растворитель в эквиобъемном количестве. В качестве референтного средства использовали трайкор (фенофибрат – современный гиполипидемический препарат) в дозе 12,5 мг/кг по схеме введения сходной с ТО и ТК. Через 1 час после последнего введения исследуемых соединений вводили внутрибрюшинно раствор адреналина в дозе 1,5 мг/кг [4]. Перед введением адреналина и через 30 минут после осуществляли забор крови из подъязычной вены. В крови определяли содержание свободного глицерина и глюкозы. Определение биохимических показателей сыворотки крови выполняли на автоматическом биохимическом анализаторе BS-380 (Mindray) с использованием стандартных наборов реактивов фирмы Diasis (Германия).
Уровень глюкозы в сыворотке животных определяли ферментативным фотометрическим тестом «GOD-PAP» с использованием глюкозооксидазы. Принцип определения: определение глюкозы ферментативным окислением в присутствии глюкозооксидазы. Окрашенный индикатор хинонимин образуется из фенола и 4-аминоантипирина под действием пероксида водорода при каталитическом воздействии пероксидазы (реакция Триндера).
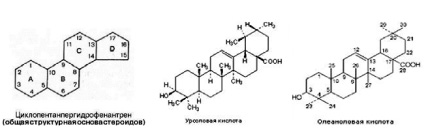
Структурные формулы: циклопентанпергидрофенантрен (общая структурная основа стероидов), урсоловая кислота, олеаноловая кислота
Уровень свободного глицерина в сыворотке животных определяли колориметрическим ферментативным тестом с использованием глицерол-3-фосфатоксидазы. Принцип определения: индикатором является хинонимин, который образуется из 4-аминоантипирина и 4-хлорфенола под действием перекиси водорода при каталитическом участии пероксидазы.
Увеличение оптической плотности при 546 нм прямо пропорционально концентрации свободного глицерина и глюкозы в образце.
Статистическая обработка проводилась с использованием t-критерия Стьюдента и U-критерия Манна – Уитни с предварительным анализом выборки на нормальность распределения. Критический уровень значимости при проверке статистических гипотез в данном исследовании принимали равным 0,05.
Результаты исследования и их обсуждение
При исследовании интенсивности липолиза в жировой ткани, индуцированного введением адреналина, выполняли определение концентрации в крови одного из конечных продуктов гидролиза триглицеридов – глицерина. О влиянии соединений на липолитические процессы в клетках жирового депо животных судили по степени прироста концентрации данного метаболита. Кроме того, дополнительно регистрировали уровень глюкозы, в регуляции которого катехоламины принимают активное участие. Парентеральное введение адреналина привело к пятикратному росту концентрации свободного глицерина в крови во всех экспериментальных группах. Данные изменения достоверно не отличались между всеми опытными группами и контрольной группой животных (табл. 1).
Как видно из данных (табл. 2), введение адреналина сопровождалось прогнозируемым ростом концентрации глюкозы в крови в большей части групп (контроль – 45 %, ТК – 28 %, ТО – 16 %), за исключением животных, получавших трайкор. В группе животных, которым вводили референтное средство, напротив, отмечено развитие гипогликемии.
Таблица 1
Изменение уровня свободного глицерина в сыворотке крови после адреналиновой стимуляции на фоне курсового введения ТО и ТК
|
Показатели Группы |
Свободный глицерин, ммоль/л |
||
|
до |
после |
Δ |
|
|
Контроль, n = 8 |
0,124 ± 0,036 |
0,516 ± 0,106▼ |
0,418 ± 0,101 |
|
ТК, 100 мг/кг, n = 8 |
0,086 ± 0,016 |
0,591 ± 0,109▼ |
0,505 ± 0,111 |
|
ТО, 100 мг/кг, n = 8 |
0,143 ± 0,028 |
0,66 ± 0,077▼ |
0,518 ± 0,072 |
|
Трайкор, 12,5 мг/кг, n = 8 |
0,139 ± 0,043 |
0,624 ± 0,065▼ |
0,485 ± 0,203 |
Примечания:
ТО – тритерпеноиды облепихи, ТК – тритерпеноиды клюквы;
* – достоверная разница по отношению к контролю, р ≤ 0,05;
▼ – достоверная разница по отношению к исходным значениям, р ≤ 0,05;
Δ – разница между значениями показателя после введения адреналина и до введения адреналина.
Таблица 2
Изменение уровня глюкозы в сыворотке крови после адреналиновой стимуляции на фоне курсового введения ТО и ТК
|
Показатели Группы |
Глюкоза, ммоль/л |
||
|
до |
после |
Δ |
|
|
Контроль, n = 8 |
4,57 ± 0,50 |
6,64 ± 1,47 |
2,07 ± 1,19 |
|
ТК, 100 мг/кг, n = 8 |
4,29 ± 0,49 |
5,48 ± 1,08 |
1,19 ± 0,42 |
|
ТО, 100 мг/кг, n = 8 |
4,77 ± 0,10 |
5,69 ± 0,83 |
0,93 ± 0,29 |
|
Трайкор, 12,5 мг/кг, n = 8 |
4,33 ± 0,03 |
3,15 ± 0,90*▼ |
–1,18 ± 0,32*▼ |
Примечания:
ТО – тритерпеноиды облепихи, ТК – тритерпеноиды клюквы;
* – достоверная разница по отношению к контролю, р ≤ 0,05;
▼ – достоверная разница по отношению к исходным значениям, р ≤ 0,05;
Δ – разница между значениями показателя после введения адреналина и до введения адреналина.
В механизмах поддержания пула липидов в крови большое значение имеет мобилизация липидов из жировых депо организма в результате активации липолитических механизмов [5], и некоторые гиполипидемические препараты (никотиновая кислота) способны подавлять выброс липидов путем подавления внутриклеточного липолиза [3]. Полученные нами результаты свидетельствуют, что на фоне приема ТО, ТК и трайкора не происходит нарушений в механизме аденилатциклазной активации липолиза, вызванной введением адреналина. В данных исследованиях установлено, что введение исследуемых субстанций и препарата сравнения сопровождается естественной активацией липолиза, причем ее уровень достоверно не отличается от изменений в контрольной группе животных. Таким образом, в данных экспериментальных условиях ТК, ТО и трайкор не оказали влияния на тканевой липолиз. Регистрация характера изменений уровня глюкозы, которые представляли интерес с позиции оценки влияния соединений на известные механизмы гуморальной регуляции уровня глюкозы в крови, которая через ряд ключевых метаболитов имеет тесную связь с обменом липидов. Было установлено, что введение ТК и ТО не оказало влияния на изменение концентрации глюкозы в крови после адреналиновой инициации гипергликемии, а на фоне введения трайкора отмечена четкая картина нарушения физиологического механизма стимуляции выхода глюкозы в кровяное русло, так как вопреки ожидаемому стандартному сдвигу концентрации глюкозы в крови в сторону увеличения после инъекции адреналина, отмечали обратную картину – достоверное по отношению к контролю снижение ее концентрации.
Выводы
1. Курсовое введение ТО и ТК не оказывает влияния на гормонально зависимый липолиз в жировых депо животных.
2. Курсовое введение ТО и ТК не оказывает влияния на механизмы поддержания уровня глюкозы в крови.
3. Курсовое введение трайкора (фенофибрата) в дозе 12,5 мг/кг вызывает нарушение физиологических механизмов регуляции уровня глюкозы в крови.
Рецензенты:
Василенко Ю.К., д.м.н., профессор кафедры биохимии и микробиологии ПМФИ филиала ГБОУ ВПО ВолгГМУ Минздрава России, г. Пятигорск;
Калашникова С.А., д.м.н., профессор кафедры морфологии и патологии ПМФИ филиала ГБОУ ВПО ВолгГМУ Минздрава России, г. Пятигорск.
Библиографическая ссылка
Горбатюк Н.О., Черников М.В., Терехов А.Ю., Маширова С.Ю. ИЗУЧЕНИЕ ВЛИЯНИЯ СУММЫ ТРИТЕРПЕНОВЫХ КИСЛОТ ОБЛЕПИХИ (Hippophaë rhamnoides L.) И КЛЮКВЫ (Vaccinium oxycoccos L.) НА ЛИПОЛИЗ В ЖИРОВОЙ ТКАНИ ЛАБОРАТОРНЫХ ЖИВОТНЫХ ПРИ КУРСОВОМ ВВЕДЕНИИ // Фундаментальные исследования. 2015. № 2-19. С. 4231-4234;URL: https://fundamental-research.ru/en/article/view?id=37935 (дата обращения: 30.05.2026).