


Scientific journal
Fundamental research
ISSN 1812-7339
"Перечень" ВАК
ИФ РИНЦ = 1,798
PROBLEMS OF BREEDING OF STURGEON LARVAE AND FRY UNDER INTENSIVE AQUACULTURE CONDITIONS AND WAYS OF THEIR SOLUTION

Интенсивное осетроводство – одно из перспективных направлений современной аквакультуры. Однако при всех положительных технологических возможностях, позволяющих контролировать и регулировать качество среды обитания и питания, проводить профилактические и лечебные мероприятия, интенсивное рыбоводство не снимает проблемы, связанные со здоровьем и, соответственно, с ростом, развитием и выживаемостью рыб. Наиболее распространенными заболеваниями осетровых в интенсивной индустриальной аквакультуре являются алиментарные, что обусловлено кормлением исключительно искусственными комбикормами. Плохое качество кормов, несоответствие их физиологической потребности рыб в питательных и биологически активных веществах, неправильный режим кормления ухудшают физиологическое состояние организма, способствуют замедлению роста и развитию тяжелых патологий, часто вызывающих массовую гибель. Более 50 % потерь приходится на дисбактериоз, в частности его специфическое проявление – тимпания, как следствие нарушения нормальной эндомикрофлоры кишечника.
В отечественной аквакультуре согласно принятым технологиям личинок и мальков осетровых рыб кормят единым стартовым комбикормом в течение 35–45 суток до выпуска в естественные водоемы или товарное выращивание [5, 7].
Для современных стартовых комбикормов характерна высокая дисперсность протеина и повышенная доля мономерных форм, что положительно сказывается на выращивании личинок. Однако длительное кормление такими кормами не учитывает поэтапное формирование ферментовыделительной системы молоди в ранний постэмбриональный период развития. Установлено, что соотношение активности между основными пищеварительными ферментами практически стабилизируется к 25 суткам активного питания мальков, что свидетельствует о новом уровне развития их организма и предполагает необходимость качественно нового состава комбикорма. Согласно нашим наблюдениям при длительном кормлении мальков высокодисперсными кормами (более 25 суток) у них зачастую наблюдается тимпания, что, соответственно, существенно снижает результативность выращивания молоди. Это обусловило задачу наших исследований, которая заключалась в оптимизации процесса кормления мальков осетровых как фактора, определяющего их продуктивность и жизнеспособность.
Материал и методы исследований
Объектом исследований служили личинки и мальки бестера. Кормление их осуществляли по следующей схеме: 1-й этап – кормление в течение 20 суток от начала активного питания кормом рецептуры Старт-1; 2-й этап – после сортировки на 2 группы в течение 21–45 суток кормление 1-й группы Стартом-1 и 2-й группы Стартом-2.
Рыб содержали в бассейнах ИЦА-1 площадью 1 м2 с круговым током воды. Термический, кислородный и гидрохимический режим (в части ионно-солевого и биогенного состава) в бассейнах был достаточно благоприятным для выращивания осетровых на протяжении всего периода исследований (t = 19,7–25,3 °С, О2 = 6,6–7,8 мг/л, рН – 8,0–8,3 ед.).
Биологическое и продуктивное действие комбикормов и физиологическое состояние молоди бестера оценивали по темпу роста, выживаемости, упитанности по Фультону, затратам кормов, эффективности использования протеина (ЭИП) и энергии (ЭИЭ) на единицу прироста. Содержание основных групп органических и минеральных веществ, фракционный состав общих липидов определяли по общепринятым методикам в прописи Н.А. Абросимовой с соавторами [3].
Жирные кислоты определяли методом газожидкостной хроматографии на хроматографе «ЦВЕТ-5». В качестве метчиков использовали стандартные смеси метиловых эфиров жирных кислот – «Sigma-189-1» и «Sigma-189-6».
Определение активности супероксиддисмутазы проводили гидроксиламиновым методом [10], a-токоферола – флуорометрическим методом [11].
Результаты исследований и их обсуждение
Известно, что на начальных этапах постэмбриогенеза у осетровых, как и других рыб, доминирует мембранное пищеварение [8, 9]. Повышенный уровень высокодисперсных белков в диетах рыб на начальных этапах кормления оказывает значительное улучшение роста личинок. В дальнейшем с возрастом наблюдается снижение темпа роста, сопровождающееся физиологическими изменениями, такими как ухудшение картины крови, увеличение печени и изменение ее структуры [8], что подтверждается теорией А.М. Уголева [9]. Согласно этой теории при длительном кормлении диетами с преобладанием мономерных форм вступает в действие бактериостимулирующий фактор, снижающий доступность таких кормов. Кроме того, для нормального развития организма необходима соответствующая нагрузка, в частности перевод организма на полимерные формы питательных веществ с более высокой молекулярной массой.
Основываясь на теории А.М. Уголева, нами были разработаны 2 варианта комбикормов, достаточно близких по содержанию основных групп органических и минеральных веществ, что достигалось соответствующим набором кормового сырья, но при этом они отличались фракционным составом протеина (табл. 1).
Оба варианта кормов обогащали дополнительно 2 % кормовой липидной добавки. Кроме того, в корм рецептуры Старт-2 вводили 0,2 % лактобактерина.
Кормление личинок бестера начинали после перехода их на активное питание при массе 33 мг согласно разработанной схеме (табл. 2).
Результаты выращивания на 1 этапе кормления сравнимы с литературными данными [4, 6]. Положительный эффект этих кормов, в том числе Старт-1, обусловлен высокой дисперсностью растворимых белковых веществ за счет гидролизатов и бульонов.
Таблица 1
Фракционный состав протеина экспериментальных кормов, г/кг
|
Вариант комбикорма |
Сумма растворимых белковых веществ |
Высокомолекулярные белки, М.М. более 10 тыс. Да |
Полипептиды, М.М. 1,5–10 тыс. Да |
Олигопептиды, М.М. менее 1,5 тыс. Да |
|
Старт-1 |
218,9 |
45,1 |
85,8 |
88,0 |
|
Старт-2 |
132,7 |
33,1 |
59,3 |
40,3 |
Таблица 2
Рыбоводно-биологические показатели выращивания молоди бестера
|
Показатели |
1 этап |
2 этап |
|
|
Старт-1 |
Старт-1 |
Старт-2 |
|
|
Начальная масса, г |
0,033 ± 0,001 |
0,624 ± 0,15 |
0,624 ± 0,15 |
|
Конечная масса, г |
0,624 ± 0,15 |
2,433 ± 0,162 |
3,221* ± 0,165 |
|
Темп роста, мг/сут |
29,55 |
72,36 |
103,88 |
|
Выживаемость, % |
62,3 |
64,5 |
83,9 |
|
Коэффициент упитанности, ед. |
0,69 ± 0,01 |
0,88 ± 0,01 |
0,87 ± 0,01 |
|
Затраты кормов, 1/г прироста |
1,2 |
1,3 |
1,1 |
|
ЭИП, % |
– |
28,4 |
37,1 |
|
ЭИЭ, % |
– |
32,2 |
41,1 |
Примечание. * – P > 0,05.
По достижении средней массы около 0,6 г мальков бестера после сортировки по массе пересаживали поровну в бассейны для дальнейшего выращивания.
Полученные рыбоводно-биологические показатели свидетельствовали о более высокой эффективности смены рациона на 2-м этапе выращивания по сравнению с традиционным методом.
Так, весовой рост мальков при смене корма Старт-1 на Старт-2 был выше на 43,6 %, что обеспечило более высокую на 32,4 % конечную массу. Смена рациона способствовала и большей на 30 % выживаемости мальков.
Известно, что наиболее оптимальным является рацион, при котором протеин максимально используется на рост, а энергетические траты осуществляются за счет липидов и углеводов пищи [9]. На 2-м этапе кормления комбикормом Старт-2 затраты кормов на единицу прироста уменьшились на 0,2 ед., а ЭИП и ЭИЭ на прирост рыб повысилась соответственно на 30,6 и 27,6 %. Положительный рыбоводный эффект подтвердился и физиологическим состоянием рыб (особи с признаками тимпании исключались).
Различия в содержании органических и минеральных веществ у мальков бестера обеих групп по завершении 2-го этапа были незначительны и не превышали 5,4 %.
Известно, что липиды являются важнейшими биологическими эффекторами, регуляторами и медиаторами, участвующими практически во всех важнейших физиологических процессах, происходящих в организме, и биохимических реакциях. Вероятно, поэтому более значимы различия липидного состава мальков (табл. 3).
Так, содержание триацилглицеридов, моноацилглицеридов и НЭЖК уменьшилось соответственно на 11,1; 12,5 и 12,7 % при одновременном повышении уровня фосфолипидов и эфиров холестерина – на 30,6 %. Различия по содержанию холестерина и диацилглицеридов были незначительными и составили соответственно 6,3 и 4,8 %.
Таблица 3
Липидный состав мальков бестера по завершении кормления, % общих липидов
|
Показатели |
1-я группа |
2-я группа |
|
Триацилглицериды |
58,6 ± 1,1 |
52,1 ± 1,1* |
|
Фосфолипиды |
20,6 ± 0,5 |
26,9 ± 0,9* |
|
Холестерин |
7,9 ± 0,2 |
8,4 ± 0,1 |
|
Эфиры холестерина |
3,6 ± 0,1 |
4,2 ± 0,3 |
|
Моноацилглицериды |
1,6 ± 0,3 |
1,4 ± 0,4* |
|
Диацилглицериды |
2,1 ± 0,2 |
2,2 ± 0,6 |
|
НЭЖК |
5,5 ± 0,4 |
4,8 ± 0,5* |
Примечание. НЭЖК – неэстерифицированные жирные кислоты; * – P > 0,05.
Отмеченные различия в составе общих липидов свидетельствуют об улучшении физиологического статуса молоди при смене рациона и подтверждаются существенным повышением уровня фосфолипидов и снижением содержания моноацилглицеридов и НЭЖК.
Адекватным показателем физиологического состояния рыб служат соотношение полиеновых кислот ωЗ/ω6, фосфолипидно-триацилглицериновый коэффициент (ФЛ/ТАГ), коэффициент Дьердии (ХС/ФЛ), уровень антиоксидантов, характеризующие липидный и энергетический обмен [1, 12]. В результате анализа отмечены различия данных показателей у мальков бестера в зависимости от состава корма (рисунок).
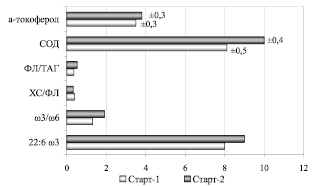
Показатели физиологического состояния молоди бестера по завершении кормления: α-токоферол, СОД – ед. активности; 22:6 ω3 – % суммы жирных кислот
Так, по завершении 2-го этапа кормления у молоди на корме Старт-2 фосфолипидно-триацилглицериновый коэффициент и соотношение w3/w6 в общих липидах превышали аналогичные показатели у рыб на корме Старт-1 на 48,6 и 46,2 % при близких величинах коэффициента Дьердии. Кроме того, соотношение w3/w6 и содержание докозагексаеновой жирной кислоты (22:6 w3) у молоди на корме Старт-2 также превышало данный показатель по сравнению со Стартом-1 соответственно на 46,2 и 12,5 %.
Полученные нами показатели липидного и обмена согласуются с данными Е.Б. Абросимовой [2], согласно которым у физиологически здоровой молоди бестера величина фосфолипидно-триацилглицеринового коэффициента и коэффициента Дьердии составляет соответственно 0,44 и 0,33 ед.
Активность СОД у бестера на корме Старт-2 была выше на 23,4 % при, хотя и незначительном, но повышении активности a-токоферола. Можно предположить, что для поддержания сбалансированного метаболизма, в том числе для регуляции процессов перекисного окисления липидов, у молоди при смене корма расходуется меньше составляющих антиоксидантную защиту организма, что имеет важное значение в адаптации организма к эндо- и экзофакторам.
Таким образом, своевременная смена рациона, основанная на возрастных изменениях функциональных особенностей пищеварительной системы ранней молоди бестера, способствовала повышению эффективности выращивания за счет повышения темпа роста, выживаемости мальков и эффективности использования протеина и энергии на рост, а также снижения затрат и эффективности использования корма на прирост. Кроме того, при такой системе кормления у молоди бестера тимпании не наблюдали, в том числе в подконтрольных производственных бассейнах. При кормлении однообразным кормом количество мальков с различной стадией тимпании составляло более 30 %.
Рецензенты:Пономарев С.В., д.б.н., профессор, директор инновационного центра «Биоаквапарк – НТЦ аквакультуры», г. Астрахань;
Шкурат Т.П., д.б.н., профессор, зав. кафедрой генетики и биохимии, Академия биологии и биотехнологии, Южный федеральный университет, г. Ростов-на-Дону.
Работа поступила в редакцию 14.01.2015.
Библиографическая ссылка
Абросимова К.С., Абросимова Н.А., Васильева Л.М. ПРОБЛЕМЫ ВЫРАЩИВАНИЯ ЛИЧИНОК И МАЛЬКОВ ОСЕТРОВЫХ РЫБ В ИНТЕНСИВНОЙ АКВАКУЛЬТУРЕ И ПУТИ ИХ РЕШЕНИЯ // Фундаментальные исследования. 2015. № 2-9. С. 1882-1886;URL: https://fundamental-research.ru/en/article/view?id=37326 (дата обращения: 19.05.2026).