


Scientific journal
Fundamental research
ISSN 1812-7339
"Перечень" ВАК
ИФ РИНЦ = 1,798
INFORMATION POSSIBILITIES OF USE OF MU AND BETA EEG RHYTHMS OF THE DOMINANT HEMISPHERE IN THE CONTRUCTION OF BRAIN-COMPUTER INTERFACE

При исследовании корковых процессных механизмов регистрируются изменения разной выраженности в ответах всех диапазонах ЭЭГ [1, 2, 6]. При планировании движений наибольшую информационную ценность представляют мю- и бета-ритмы ЭЭГ [7, 15]. А при высокой активации проприоцептивной системы высокое значение имеют тета- и дельта-ритмы ЭЭГ [2]. В настоящей работе внимание сосредоточено на изучении ЭЭГ-коррелятов планирования и начала движения – в мю- и бета-частотных диапазонах ЭЭГ.
Частота мю-ритма определяется частотными диапазонами ЭЭГ в 8–10 и 10–12 Гц [11]. Некоторые компоненты бета-ритмов частотой 13–25 Гц являются гармониками мю-ритма [12]. В состоянии покоя максимальная мощность мю-ритма отмечается в соматосенсорной коре [4]. Важнейшая особенность мю-ритма состоит в том, что он изменяется только под влиянием проприоцептивных раздражителей и не реагирует или мало реагирует на воздействие других, например, световых или звуковых сигналов. В отличие от классического альфа-ритма мю-ритм асимметричен и не проявляет двусторонней когерентности между полушариями [14]. Известно, что десинхронизация сенсомоторных мю- и бета-ритмов в моторных и сенсомоторных зонах контралатерального моторному акту большого полушария головного мозга происходит при реальных движениях и при их мысленном представлении [7]. Кроме этого, для удобства пользователя нейрокомпьютерного интерфейса актуальным является уменьшение количества отведений при регистрации сигналов мозга на ЭЭГ. Поэтому целью настоящей работы было исследование мю- и бета-ритмов ЭЭГ в доминантном левом полушарии при реальных и воображаемых движениях.
Материал и методы исследования
В работе приняли участие 7 испытуемых-правшей в возрасте 18–19 лет, давших информированное согласие на участие в исследовании. ЭЭГ регистрировалась монополярно с помощью системы «Нейрон – Спектр – 4/ВПМ». 17 электродов ЭЭГ были высокоплотно локализованы по системе 10–10 в проекциях первичных, вторичных моторных и соматосенсорных зон левого полушария ЭЭГ. Частота квантования ЭЭГ составляла 200 Гц, пределы фильтрации от 0 до 70 Гц, чувствительность 5 мкВ. Использовался Notch фильтр для подавления наводки электричества бытовой сети. Импеданс всех каналов был ниже 20 кОм. Во время записи ЭЭГ испытуемые находились в темной звукоизолированной комнате с закрытыми глазами в положении сидя. ЭЭГ у испытуемых регистрировалась в следующих режимах: ЭЭГ фоновая; ЭЭГ при реальном выполнении движений (5 сессий); ЭЭГ при воображении движений (5 сессий). Продолжительность одной сессии – 10 с. Испытуемыми выполнялись реальные и воображаемые движения: сгибание пальцев правой руки; тыльное сгибание правой стопы. Полученные данные обрабатывались статистически с помощью системы StatSoft Statistica 6.0.
Результаты исследования и их обсуждение
При анализе всего спектра ЭЭГ без дифференцировки на частотные составляющие в проекциях моторных и сенсомоторных зон коры левой гемисферы при реальных и воображаемых движениях конечностей не выявлено статистически значимой разницы значений мощности ритмов ЭЭГ относительно фона (рисунок, табл. 1, 2).
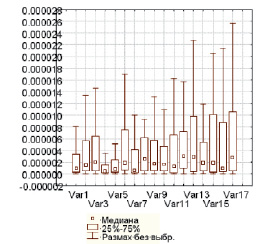
Диаграммы размаха мощности всего спектра ЭЭГ по 17 отведениям при реальном движении руки. По горизонтали – номера отведений, по вертикали – значения мощности (мкВ2)
Таблица 1
Средние значения мощности ритмов ЭЭГ при реальном и воображаемом сгибании пальцев правой руки, мкВ 2 (М ± m)
|
Ритмы |
Фон |
Движение |
Р |
|
|
Реальное |
Воображаемое |
|||
|
Мю- |
1,2666Е-5 ± 2,956Е-6 |
1,0132Е-5 ± 0,9829Е-6 |
1,1125Е-5 ± 1,0029Е-6 |
0,005 |
|
Бета- |
4,2772Е-7 ± 7,2361Е-8 |
1,4171Е-7 ± 2,2436Е-8 |
1,521Е-7 ± 4,2436Е-8 |
0,001 |
|
Весь спектр |
2,407Е-6 ± 0,5479Е-6 |
2,6041Е-6 ± 1,4029Е-6 |
2,4705Е-6 ± 1,0329Е-6 |
0,4 |
Таблица 2
Средние значения мощности ритмов ЭЭГ при реальном и воображаемом тыльном сгибании стопы, мкВ2 (М ± m)
|
Ритмы |
Фон |
Движение |
Р |
|
|
Реальное |
Воображаемое |
|||
|
Мю- |
1,3566Е-5 ± 1,478Е-6 |
1,0853Е-5 ± 0,7829Е-6 |
1,1235Е-5 ± 1,1029Е-6 |
0,005 |
|
Бета- |
4,7472Е-7 ± 7,2521Е-8 |
1,4771Е-7 ± 2,2576Е-8 |
1,521Е-7 ± 4,2436Е-8 |
0,001 |
|
Весь спектр |
2,397Е-6 ± 0,5479Е-6 |
2,6471Е-6 ± 1,2329Е-6 |
2,4505Е-6 ± 1,3829Е-6 |
0,3 |
Статистически достоверное уменьшение (десинхронизация) значений мощности ритмов ЭЭГ выявлено только в узких частотных спектрах, в мю- (11–13 Гц) и бета- (14–30 Гц), как при реальных, так и во время воображаемых движений (табл. 1, 2).
Однако не было достоверного различия значений мощности мю- и бета-ритмов ЭЭГ при реальных и воображаемых движениях. Как при реальных, так и при воображаемых движениях кисти и стопы мощность мю-ритма уменьшалась на 80 %. Продолжительность десинхронизации мю-ритма составляла 3–4 с. В течение последующих 2 с мощность в мю-частотном диапазоне ЭЭГ увеличивалась на 50 % от предыдущего минимального значения (десинхронизация сменялась на синхронизацию). При реальных и воображаемых движениях был отмечен аналогичный характер изменения мощности в бета-частотном диапазоне ЭЭГ. В этих условиях исследования мощность бета-ритма уменьшалась в 3,13–3,2 раза по сравнению с фоном (табл. 1, 2). Продолжительность десинхронизации бета-ритма составляла 1–2 с.
Десинхронизация мю-ритма при планировании движения объясняется активацией сенсомоторной области с последующей выраженной синхронизацией этого ритма на начальной стадии выполнения движения [13]. Этот процесс более выражен в контралатеральной гемисфере [5]. Феномен синхронизации мю-ритма ЭЭГ во время выполнения движения связывают с процессами двусторонней дезактивации сенсомоторных зон или сбросом информации в моторные центры [8, 9, 12]. В роли модулирующего паттерна по отношению к мю-ритму ЭЭГ рассматривают систему зеркальных нейронов [3].
Наши данные совпадают с результатами других авторов, проводивших исследования с билатеральной регистрацией ЭЭГ. Выявленная нами сходная динамика изменений мощности мю- и бета-ритмов ЭЭГ в условиях унилатеральной локализации электродов при реальных и воображаемых движениях может быть основой для детекции планирования движения кисти или стопы и использования подобного приема в нейрокомпьютерных интерфейсах.
Заключение
В работе показана возможность использования двух асимметричных информационных каналов в виде уменьшения мощности мю- и бета-ритмов ЭЭГ в доминантном полушарии при воображении движений верхних и/или нижних конечностей для конструирования управляемых сигналами мозга человека роботизированных систем.
Работа поддержана грантом № 4/1-ИПП. 18.06.2014 г. ИФ Самарской области.
Рецензенты:Мирошниченко И.В., д.м.н., профессор, заведующий кафедрой нормальной физиологии, ГБОУ ВПО «Оренбургская государственная медицинская академия» Министерства здравоохранения Российской Федерации, г. Оренбург;
Ведясова О.А., д.б.н., профессор кафедры физиологии человека и животных, ФГОУ ВПО «Самарский государственный университет», г. Самара.
Работа поступила в редакцию 02.03.2015.
Библиографическая ссылка
Пятин В.Ф., Колсанов А.В., Сергеева М.С., Захаров А.В., Антипов О.И., Коровина Е.С., Тюрин Н.Л., Глазкова Е.Н. ИНФОРМАЦИОННЫЕ ВОЗМОЖНОСТИ ИСПОЛЬЗОВАНИЯ МЮ- И БЕТА-РИТМОВ ЭЭГ ДОМИНАНТНОГО ПОЛУШАРИЯ В КОНСТРУИРОВАНИИ НЕЙРОКОМПЬЮТЕРНОГО ИНТЕРФЕЙСА // Фундаментальные исследования. 2015. № 2-5. С. 975-978;URL: https://fundamental-research.ru/en/article/view?id=36967 (дата обращения: 06.07.2026).