


Scientific journal
Fundamental research
ISSN 1812-7339
"Перечень" ВАК
ИФ РИНЦ = 1,798
TRIPEPTIDE MODULATE THE CONDITIONALLY-REFLECTIVE ACTIVITY IN A HONEY-BEE APIS MELLIFERA L.

В конце ХХ в. было показано модулирующее влияние нейропептидов в интегративной деятельности нервной системы у позвоночных и беспозвоночных животных. Нейропептиды принимают участие в запоминании, обучении, механизме сна. Таким образом, нейропептиды являются классом универсальных химических регуляторов, значимость которых «простирается от влияния на функции отдельных групп клеток до управления работой целых систем и органов, включая сложные акты поведения» [1]. В настоящее время открыто около 100 нейропептидов, которые синтезируются различными типами нейронов головного мозга. Большинство нейропептидов синтезируется в виде предшественников и переходят в активную форму путем органиченного протеолиза. Этот биологический феномен лег в основу создания нейропротекторных лекарств нового поколения – коротких пептидов, представляющих собой активные центры полипептидных молекул. Так, пептиды семакс и кортаген успешно применяются в лечении дисфункций центральной нервной системы различного генеза [3]. В Санкт-Петербургском институте биорегуляции и геронтологии был сконструирован пептид EDR (пинеалон, Lys-Glu-Arg) [9]. Нейропротекторные свойства пептида EDR были изучены в культурах клеток, в опытах на животных и в клинической практике [3, 4, 8].
Целью работы явилось изучение влияния пептид EDR на функцию высших отделов нервной системы, обеспечивающих когнитивную деятельность, в частности формирование памяти. Биохимическая и физиологическая общность механизмов формирования памятного следа в нервной системе у позвоночных и беспозвоночных животных позволяет использовать в качестве модельного объекта насекомых.
Исследование проведено на модельном объекте – медоносной пчеле краинской расы Apis mellifera carnica Pollm. Известно, что способность к ассоциативному обучению наибольшей сложности достигает именно у общественных насекомых.
Многолетние исследования свидетельствуют о способности этого вида насекомых к осуществлению высших форм нервной деятельности: к ассоциативному обучению, к долговременному хранению в памяти приобретаемого в процессе пищедобывательной летной деятельности индивидуального опыта, об использовании при этом рецепторных, сигнальных, трансдукционных механизмов, сходных с млекопитающими. В настоящее время ассоциативное обучение у медоносной пчелы изучено на поведенческом, нейрональном, межклеточном и клеточном уровнях, включая изучение генетического аппарата. В модели условно-рефлекторной деятельности пчелы Apis mellifera исследована роль глутаматных рецепторов в формировании памяти, изучены некоторые нейропротекторные свойства аминокислот и пептидов, проведены сравнительные генетические исследования функции ЦНС с другими видами модельных насекомых, например с Drosophila melanogaster [1, 2].
Материалы и методы исследования
Объектом исследования служила медоносная пчела краинской расы Apis mellifera carnica Pollm в возрасте 10–40 дней. Для изучения условно-рефлекторной деятельности пчел оценивали образование пищевого условного рефлекса вытягивания хоботка на обонятельный раздражитель (PER – Proboscis Extension Response conditioning). У фиксированных за крылья пчел вырабатывали условный рефлекс вытягивания хоботка путем однократного сочетания запаха гвоздики и пищевого подкрепления – 50 % раствора сахарозы. Через 1 мин (кратковременная память) и 180 мин (долговременная память) по окончании процедуры обучения у пчел проверяли наличие условной реакции на запах. До обучения у пчел оценивали сенсорную возбудимость – наличие спонтанной реакции вытягивания хоботка на еще неподкрепленный запах (при наличии таковой пчел выбраковывали) и пищевую возбудимость – вытягивание хоботка в ответ на соприкосновение раствора сахарозы с антеннами (вкусовая рецепция) (в отсутствие таковой пчел выбраковывали). За 3 ч до процедуры обучения пчел изолировали, лишая их пищи и контакта с семьей, для повышения пищевой мотивации. Фоновый условно-рефлекторный уровень определяли как количество пчел, сохраняющих в памяти условную реакцию в контрольной группе (инъекции физиологического раствора) и выражали в %. За 30 мин до обучения пчелам дорзально в торакс делали инъекции 2 мкл раствора пептида EDR в ряду концентраций 10-11М–10-4М (опыт) или 2 мкл физиологического раствора (контроль). В каждой серии экспериментов использовали 60 пчел. Число пчел, ответивших условной реакцией, оценивали в % по отношению к контрольному уровню (принятому за 100 %).
Статистическая обработка данных включала в себя подсчет среднего арифметического, стандартного отклонения от среднего и доверительного интервала для каждой выборки и проводилась в программе «Statistica 7.0». Для анализа вида распределения и проверки нулевой гипотезы использовали критерий Шапиро-Уилка. Для оценки статистической однородности нескольких выборок были использованы непараметрические процедуры однофакторного дисперсионного анализа (критерий Крускала–Уоллиса). Для статистического анализа достоверных различий характеристик памяти у пчел применяли двухвыборочный критерий Вилкоксона ранговых сумм. Различия между группами считали статистически значимыми при р < 0,05.
Результаты исследования и их обсуждение
Эффективные концентрации пептида EDR находились в диапазоне 10-10–10-5 М. Если пептид EDR оказывал стимулирующее или ингибирующее влияние на кратковременную память при использовании всех указанных концентраций, то воздействие на долговременную память оказывали только низкие концентрации пептида (10-10–10-8 М). Стимулирующий или ингибирующий эффект пептида EDR зависит от фонового условно-рефлекторного уровня. В среднем 67,8 ± 1,3 % пчел сохраняют в кратковременной/долговременной памяти выработанный условный рефлекс на обонятельный раздражитель при пищевом подкреплении. В данном исследовании все насекомые были разделены на 2 подгруппы – в первой условно-рефлекторный фон отклонялся от среднего значения в меньшую сторону, когда 47,6 ± 3,5 % пчел сохраняли в памяти условную реакцию. Во второй подгруппе контрольный условно-рефлекторный фон отклонялся от среднего значения в большую сторону – 92,3 ± 4,7 % пчел сохраняли в памяти условную реакцию. Введение пептида EDR в случае исходного низкого условно-рефлекторного уровня оказывало стимулирующее воздействие на процессы, способствующие сохранению в памяти выработанного условного рефлекса (рис. 1). При концентрации пептида 10-8 М количество пчел, сохраняющих в кратковременной памяти условную реакцию, увеличивалось на 23 % по сравнению с начальным низким условно-рефлекторным уровнем. При концентрации пептида 10-7 М количество таких пчел увеличивалось на 40 %, при концентрации 10-6 М – на 28 %, при концентрации 10-5 М – на 36 %. На сохранение в долговременной памяти выработанного рефлекса у пчел с исходно низким условно-рефлекторным уровнем пептид EDR не оказывал влияния.
Иная картина наблюдалась при исходном высоком фоновом условно-рефлекторном уровне (рис. 2). В этом случае при воздействии пептида EDR в диапазоне 10-10–10-7 М количества пчел, сохраняющих в кратковременной и долговременной памяти условную реакцию, значительно снижалось. При введении пептида в концентрации 10-10 М количество пчел, сохраняющих в кратковременной памяти условную реакцию, уменьшалось на 34 %, а количество пчел, сохраняющих в долговременной памяти условную реакцию, снижалось на 15 % по сравнению с исходным высоким условно-рефлекторным уровнем. Концентрация пептида 10-9 М оказывала ингибирующее влияние только на кратковременную память. При этом количество пчел, сохраняющих в кратковременной памяти условную реакцию, уменьшалось на 38 %. Концентрация пептида 10-8 М оказывала влияние на оба вида памяти. Количество пчел, способных сохранять приобретенный опыт в кратковременной памяти, уменьшалось на 35 %, а в долговременной памяти – на 17 %. Воздействие пептида EDR в концентрации 10-7 М ингибировало долговременную память – количество пчел, сохраняющих в памяти условную реакцию, уменьшалось на 30 %.
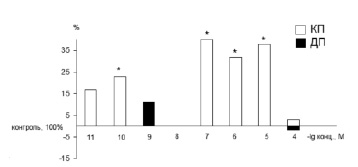
Рис. 1. Влияние пептида EDR на формирование кратковременной/долговременной памяти у медоносных пчел с исходно низкой способностью сохранять в памяти выработанный условный рефлекс. Условные обозначения (здесь и на рис. 2): белые столбики – кратковременная память, черные столбики – долговременная память по отношению к контрольному уровню (инъекции физиологического раствора), принятому за 100 %. Примечание. * – p < 0,05– по сравнению с соответствующим контролем
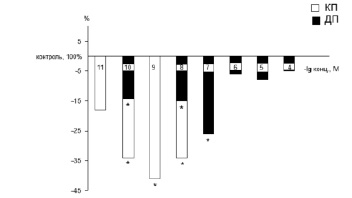
Рис. 2. Влияние пептида EDR на формирование кратковременной/долговременной памяти у медоносных пчел с исходно высокой способностью сохранять в памяти выработанный условный рефлекс
Одна и та же концентрация пептида EDR стимулировала процесс формирования способности к сохранению памяти у пчел с исходно низкой пищевой мотивацией, и ингибировала этот процесс у пчел с исходно высокой пищевой мотивацией. Таким образом, пептид EDR обладает модулирующей способностью в зависимости от уровня исходного условно-рефлекторного фона.
Проведенные исследования позволяют сделать вывод о влиянии пептида EDR на функцию высших отделов центральной нервной системы насекомых, ответственных за когнитивную деятельность. У пчелы – это парные грибовидные тела, выполняющие основную роль в обонятельном обучении и памяти при пищевом подкреплении. При исследовании нейропептидов, экспрессирующихся и утилизируемых нейронами грибовидных тел отмечаются особенности эволюции пептидергической сигнальной системы у насекомых [7, 10]. Наиболее близким по аминокислотному составу к пептиду EDR нейропептидом у пчел является оркокинин, включающий в себя глутамат (Е), аспартат (D), аргинин (R) [10]. Оркокинин регулирует циркадианные ритмы локомоторной активности насекомых, участвует в процессах нейромодуляции, регулирует синтез некоторых нейрогормонов [6, 7, 10].
Установлено, что молекулярный механизм биологической активности пептида EDR связан с его способностью регулировать экспрессию фермента 5-триптофангидроксилазы, селективно связываясь с промоторной зоной соответствующего гена по последовательности CCTGCC [4]. Известно, что 5-триптофангидроксилаза является ключевым ферментом, участвующим в синтезе серотонина. Кроме того, пептид EDR регулирует синтез серотонина в клетках нейронов млекопитающих [4]. Таким образом, пептид EDR, подобно оркокинину, способен регулировать синтез нейрогормонов.
Как уже упоминалось, оркокинин регулирует локомоторную активность у насекомых. Подобный эффект наблюдался под действием пептида EDR у людей. Пептид EDR повышал уровень энергетического обеспечения мышечной ткани, что коррелировало с увеличением экспрессии генов PPARA и PPARG, кодирующих белки, увеличивающие окислительную способность скелетных мышц [3]. Кроме того, установлено, что пептид EDR способствовал повышению умственной и физической работоспособности людей старших возрастных групп [4]. Сопоставляя эти данные с полученным модулирующим действием пептида EDR на долговременную и кратковременную память у медоносной пчелы, можно предположить эволюционную общность механизмов пептидной регуляции у млекопитающих и насекомых. Вероятно, пептидная регуляция функций нейронов является одним из древнейших механизмов нейропластичности.
Рецензенты:Медведев Д.С., д.м.н, доцент, заведующий кафедрой геронтологии и гериатрии ЧОУ ВПО «Санкт-Петербургский медико-социальный институт», г. Санкт-Петербург;
Евгений И.Ч., д.б.н., профессор, профессор кафедры биологии, эмбриологии и гистологии Санкт-Петербургской государственной академии ветеринарной медицины, г. Санкт-Петербург.
Работа поступила в редакцию 12.02.2015.
Библиографическая ссылка
Хавинсон В.Х., Лопатина Н.Г., Чалисова Н.И., Зачепило Т.Г., Линькова Н.С., Халимов Р.И., Камышев Н.Г. ТРИПЕПТИД МОДУЛИРУЕТ УСЛОВНО-РЕФЛЕКТОРНУЮ ДЕЯТЕЛЬНОСТЬ МЕДОНОСНОЙ ПЧЕЛЫ APIS MELLIFERA L. // Фундаментальные исследования. 2015. № 2-3. С. 492-496;URL: https://fundamental-research.ru/en/article/view?id=36840 (дата обращения: 09.07.2026).