


Scientific journal
Fundamental research
ISSN 1812-7339
"Перечень" ВАК
ИФ РИНЦ = 1,798
MACROPHAGOCYTES WITHIN RATS’ WOMB BODY BIOAMIC PROVISION SYSTEM DURING SEX CYCLE

Биогенные амины, обладающие широким спектром биологических эффектов [2], представляют собой важное звено в системе нейрогуморальной регуляции функций матки [1, 7]. В макрофагических клетках матки крыс установлено наличие серотонина, катехоламинов и гистамина [3, 4]. В то же время отсутствуют данные о динамике содержания указанных биоаминов в различные фазы полового цикла.
Цель работы: дифференцировать содержание катехоламинов, серотонина и гистамина в макрофагических клетках различных оболочек тела и шейки матки крыс в течение полового цикла
Материал и методы исследования
Работа выполнена на 120 интактных самках крыс репродуктивного возраста в осенне-зимний период, которые были распределены на группы по стадиям полового цикла: ранний эструс, поздний эструс, метаэструс, ранний диэструс, поздний диэструс, проэструс. Животных выводили из опыта в соответствии с «Правилами проведения работ с использованием экспериментальных животных» (приказ Минвуза от 13.11.1984 г № 724). Срезы тела матки толщиной 20 мкм изготавливались на криостате.
Методы исследования: флуоресцентно – гистохимические методы Фалька-Хилларпа в модификации для выявления катехоламинов и серотонина, метод Кросса-Эвана-Роста для дифференцировки гистамина [2]. Для выявления фагоцитирующих клеток прижизненно вводили подкожно 3 мл 1%-ного раствора трипанового синего [5].
Концентрацию биоаминов определяли в условных единицах шкалы регистратора с помощью люминесцентного микроскопа ЛЮМАМ-И3 с фотометрической насадкой ФМЭЛ-1А. Статистический компьютерный анализ осуществляли с помощью электронных таблиц Excel. Достоверность различий при сравнении величин определялась с помощью критерия Стьюдента. Для выявления и анализа сопряжений изменения параметров в динамике полового цикла применялись линейный корреляционный анализ Пирсона (r) и ранговый корреляционный анализ Спирмена (R). Критический уровень значимости при проверке статистических гипотез принимался равным 0,05.
Результаты исследования и их обсуждение
Макрофагические клетки, флуоресцирующие желтовато-зеленым цветом, располагаются в основном в эндометрии единично или небольшими скоплениями. Они содержат нередко сливающиеся гранулы разной величины и интенсивности флуоресценции. У макрофагов, цитоплазма которых плотно заполнена гранулами вводимой краски, флуоресценция слабая или отсутствует. В эндометрии некоторые из макрофагов располагаются под эпителием или непосредственно в нем, единичные - в составе содержимого просвета матки.
 а
а
 б
б
 в
в
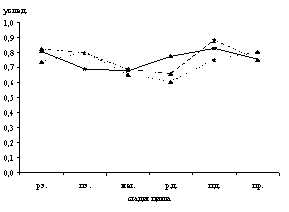
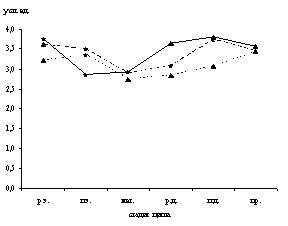
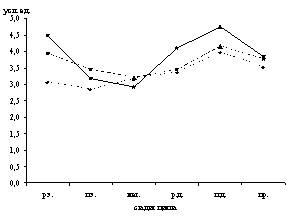
Рис. 1. Динамика содержания катехоламинов (а), серотонина (б), гистамина (в) в макрофагах тела матки крыс в процессе полового цикла
р.э. – ранний эструс, п.э. – поздний эструс, мет. – метаэструс, р.д. – ранний диэструс, п.д. – поздний диэструс, пр. – проэструс
![]()
![]() - достоверность и
- достоверность и ![]() – недостоверность отличий данного оценочного параметра от такового в предыдущей стадии полового цикла (p < 0,05)
– недостоверность отличий данного оценочного параметра от такового в предыдущей стадии полового цикла (p < 0,05)
Микроспектрофлуориметрически в макрофагах дифференцированы серотонин, гистамин и катехоламины. Максимальное содержание моноаминов в макрофагах эндометрия и миометрия зарегистрировано в поздний диэструс, а в периметрии – в проэструс (рис. 1).
В эти же стадии во всех оболочках матки отмечается наименьшая плотность пространственного распределения макрофагов (рис. 2).
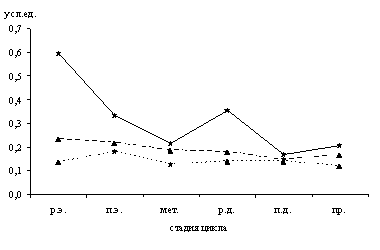
Рис. 2. Плотность пространственного распределения макрофагов тела матки крыс в процессе полового цикла
р.э. – ранний эструс, п.э. – поздний эструс, мет. – метаэструс, р.д. – ранний диэструс, п.д. – поздний диэструс, пр. – проэструс
![]()
![]() - достоверность и
- достоверность и ![]() – недостоверность отличий данного оценочного параметра от такового в предыдущей стадии полового цикла (p < 0,05)
– недостоверность отличий данного оценочного параметра от такового в предыдущей стадии полового цикла (p < 0,05)
Низкий уровень биоаминов в макрофагических клетках эндометрия выявлен в поздний эструс и метаэструс, в миометрии и периметрии – в метаэструс и ранний диэструс (рис. 1).
На протяжении всего полового цикла сохраняется высокая степень тесноты положительной линейной связи между количеством серотонина и катехоламинов в макрофагах по точкам зондирования (r = 0,70-0,98). Ранговый корреляционный анализ выявляет в эндометрии и миометрии высокую степень положительного хроносопряжения изменений в течение цикла содержания в макрофагах гистамина, серотонина и катехоламинов (R=0,714-1,0). В периметрии взаимосвязь выявлена только между изменениями уровней серотонина и катехоламинов (R=0,886).
Большое содержание моноаминов в макрофагах, по всей видимости, отражает количество их свободных форм в межклеточном пространстве, которые могут быть подвергнуты поглощению макрофагами [2].
В последнее время установлена способность макрофагов продуцировать гистамин и серотонин [8, 9].
Согласно предложенной нами рабочей гипотезе о функциональной кооперации элементов внутриорганного комплекса биоаминового обеспечения макрофаги осуществляют поглощение, накопление и, возможно, инактивацию излишков моноаминов, которые в силу тех или иных обстоятельств не утилизировались рабочими структурами [5]. Вероятно, в ряде случаев макрофаги выполняют функцию переносчиков нейромедиаторов между различными участками органа, а в некоторых случаях могут служить дополнительными их источниками.
СПИСОК ЛИТЕРАТУРЫ:
1. Абрамченко В.В., Капленко О.В. Адренергические средства в акушерской практике. СПб., ТОО ТК «Петрополис», 2000. 272 с.
2. Гордон Д.С., Сергеева В.Е., Зеленова И.Г. Нейромедиаторы лимфоидных органов. Л.: Наука, 1982. 128 с.
3. Гордон Д.С., Гунин А.Г. //Архив АГЭ. - 1988. - № 12. - С. 66.
4. Гунин А.Г., Гордон Д.С., Семенов В.Д. //Проблемы эндокринологии. – 1991. - Т. 37. № 4. - С. 49.
5. Диндяев С.В., Погорелов Ю.В. //Успехи физиол. наук. - 1993. - Т. 24. - № 4. - С. 71.
6. Диндяев С.В. Клиническая морфология яичников. Иваново, 2005. 145 с.
7. Шорников А.И., Меркулова Л.М., Ланцова Н.Н., Иванова З.И., Мочалова А.А. //Морфология. - 2006. - Т. 129. № 2. - С. 104.
8. Martins E.Jr., Ferreira A.C., Skorupa A.L. et al. //J. Leukoc. Biol. - 2004. - V. 75. № 6. - P. 1116.
9. Nakano K., Takamatsu S. //Nippon Yakurigaku Zasshi. - 2001. - V. 118. № 1. - P. 15.
Библиографическая ссылка
Диндяев С.В МАКРОФАГИ В СИСТЕМЕ БИОАМИНОВОГО ОБЕСПЕЧЕНИЯ ТЕЛА МАТКИ КРЫС В ТЕЧЕНИЕ ПОЛОВОГО ЦИКЛА // Фундаментальные исследования. 2007. № 11. С. 16-19;URL: https://fundamental-research.ru/en/article/view?id=3670 (дата обращения: 04.07.2026).