


Scientific journal
Fundamental research
ISSN 1812-7339
"Перечень" ВАК
ИФ РИНЦ = 1,798
QUANTITATIVE ANALYSIS OF THE ACTIVATION OF SIGNALING PATHWAYS OF RADIORESISTAND AND RADIOSENSITIVE CANCER CELL LINES

Проблема радиорезистентности раковых опухолей, несмотря на накопленный объем знаний, о механизмах развития противоопухолевых эффектов, остается одной из основных для онкологии. Известно, что индукция первичных изменений, вызывающих резистентность опухолей к повреждающим воздействиям, происходит, как на молекулярном, так и на клеточном уровне: взаимодействие с мишенями, блокирование отдельных внутриклеточных процессов, нарушение контроля клеточного цикла, апоптоза [1]. Не исключено участие данных изменений и на более высоких уровнях организации живой материи (межклеточные взаимодействия, ткань, системы органов, организм) [4]. Радиорезистентность опухолей – это явление, в основе которого лежит ряд молекулярных изменений, что в свою очередь формирует чувствительность раковых клеток к радиотерапии. Специфические и типовые молекулярные процессы, которые определяют ответ клетки на стресс, в том числе на токсическое или радиационное воздействие, связаны с широким диапазоном реакций – от ограничения накопления химиопрепарата внутри клетки до отмены программы гибели клетки, индуцируемой повреждающим воздействием [5]. Устойчивость раковых клеток формируется под влиянием стрессовых воздействий, в том числе в результате применения радиотерапии. Резистентность формируется на разных «ступенях» воздействия на клетку, причем на каждой «ступени» задействованы различные молекулы [6, 13]. Известно, что после MDR-трансфекции клоны лейкозной клеточной линии К-562 обнаружили повышенную устойчивость к радиации и что под влиянием ионизирующего излучения на опухолевые клетки различного гистогенеза экспрессия P-gp, MRP1 и LRP может усилиться. Поэтому причинами формирования радиорезистентности раковых опухолей являются изменение экспрессии генов и степень активации сигнальных путей, в которых они участвуют.
В большинстве транскриптомных исследований основной задачей являлся поиск генов которые дифференциально экспрессируются в экспериментальной и контрольной группе. После выявления этих генов, обычно делалось заключение о том, что тот или иной дифференциально-экспрессирующийся ген может принимать участие в формировании адаптивного ответа к экспериментальному воздействию [12]. Аналогичные работы можно найти и при исследовании радиорезистентности раковых опухолей [16]. В подобных исследованиях не учитывается то, что гены и их транскрипты являются частью сигнальных путей, состоящих из десятков и сотен взаимодействующих единиц. Кроме этого, гены, кодирующие сигнальные белки включены во множество сигнальных путей [15]. Для учета этих факторов необходимо оценивать количественную активацию сигнальных путей, компонентами которых являются дифференциально экспрессирующиеся гены. Недавно был разработан метод Oncofinder, который позволяет оценить количественно активацию сигнальных путей [7].
Цель настоящей работы – поиск генов, экспрессия которых имеет одинаковую динамику после радиационного воздействия в радиорезистентной и радиочувствительной раковых клеточных линиях и анализ количественной активации сигнальных путей, участниками которых являются эти гены.
Материал и методы исследования
Использовали радиорезистентную (К562) раковую клеточную линию хронического миелолейкоза и клеточную линию рака прямой кишки человека HCT-116 с нормальным геном ТР53 – НСТ116р. Клетки облучали рентгеновским излучением, генерируемым терапевтическим акселератором Cliniac 600 при комнатной температуре в дозах 4 Грэй одноразово. Клетки облучались в 24 луночных планшетах (объём лунки 2,5 мл). Профили экспрессии генов в клетках К562 и НСТ116р, облученных в дозе 4 Гр, изучали через 1, 12 и 24 часа после облучения с использованием микроматрицы Affymetrix серии HGU133А.
РНК выделяли из 3∙106 клеток с использованием набора для выделения РНК в соответствии с инструкцией производителя. Целостность выделенной РНК проверяли с использованием биоанализатора Agilent 2100 по целостности 18S и 28S рибосомальной. Библиотеку клонированных ДНК готовили с использованием набора GeneChipExpression 3’-Amplification One-CyclecDNASynthesisKit. Мечение биотином анти-смысловых библиотек клонированных РНК и очистка были проведены с использованием набора GeneChipExpression 3’-Amplification Reagentsfor IVT Labeling в соответствии с протоколом производителя. Количество полученной РНК и ДНК оценивалось спектрофотометрически с использованием спектрофотометра NanoDrop. Фрагментацию кРНК проводили при 94°C в термоциклере в течение 35 минут. Синтезированные биотинилированные кРНК вначале гибридизировали с контрольной матрицей «Test-3» с целью оценки качества полученных кРНК. Если качество биотинилированных кРНК соответствовало расчётному, то тогда проводили гибридизацию с матрицей HGU133А. Матрицу окрашивали стрептовидин-фикоэритрином. Окрашенную матрицу отмывали от несвязавшегося белка и сканировали на сканере GeneArray G2500A.
Отбор генов, чья экспрессия увеличивалась или уменьшалась по отношению к контролю на протяжении всего эксперимента, проводился с использованием программы Gene Selector (программу можно получить у авторов статьи), достоверность отличий задавалась на уровне p-Value ≤ 0,05.
Определение сигнальных путей, к которым относятся отобранные гены, проводилось с использованием программы PANTHER [2], базы данных Qiagen SABiosciences [3]. Количественную активацию сигнальных путей выполняли с использованием метода Oncofinder [7].
Результаты исследования
и их обсуждение
На рис. 1, а показано, что в радиорезистентной (K562) раковой клеточной линии 218 генов характеризуются повышенной экспрессией на протяжении всего эксперимента, а в радиочувствительной (HCT116p) раковой клеточной линии 141 сверхэкспрессирующихся генов. Общими для них являются 11 генов (CROCCP2, CDK1, GRB10, SLC2A3, PSRC1, SLC2A3, FANCE, MARS, NDC80, NAA35, CAP1). На рис. 1, б в радиорезистентной (K562) раковой клеточной линии найдено 149 генов, а в радиочувствительной (HCT116p) раковой клеточной линии 147 генов, чья экспрессия снижается и достигает своего минимального значения после 24 часов эксперимента. Среди них 3 гена (PRSS3, CREBL2, VPS26A), которые являются общими для радиорезистентной (K562) и радиочувствительной (HCT116p) клеточных линий. Таким образом, повышенная экспрессия одних и сниженная экспрессия других генов в радиорезистентной (K562) раковой клеточной линии может быть связана с мутациями в этих генах. Следствием этого является формирование высокой радиорезистентности раковых клеток.
Количественный анализ экспрессии генов позволяет получить новую информацию о молекулярной природе опухоли. Однако данные профилей экспрессии генов, зачастую являются противоречивыми, т.к. при обработке результатов учитывается экспрессия генов самих по себе, а не в контексте биологических процессов, за которые они отвечают. Так, все гены можно разделить на ансамбли – внутриклеточные сигнальные пути [7]. Сети сигнальных путей в клетке многочисленны и сложны. Они способны взаимодействовать и влиять друг на друга. Изменение одного компонента сигнального пути, например сверхэкспрессия гена, способно нарушить функционирование всей системы и стать причиной формирования радиорезистентности. Например, сверхэкспрессия EGFR часто встречается в опухолях человека. В большом числе исследований подчеркивается связь между EGFR и устойчивостью опухоли к ионизирующим излучениям. Huamani и соавт. продемонстрировали, что раковые клетки предстательной железы (DU145 и PC-3) экспрессируют EGFR [8, 9, 14].
Более того, в преклинических исследованиях была обнаружена корреляция экспрессии EGFR с пролиферацией, ангиогенезом и миграцией опухолевых клеток, а также с радиорезистентностью опухоли [10, 11].
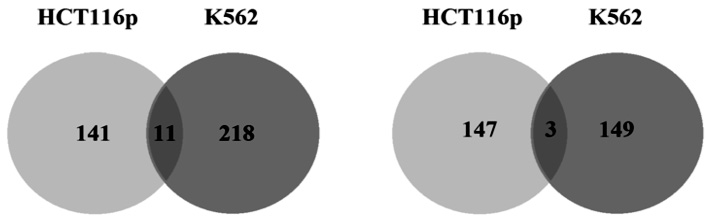
а б
Рис. 1. Количество общих генов в радиорезистентной (K562)
и радиочувствительной раковых клеточных линиях (HCT116p):
а – количество общих генов, чья экспрессия растет на протяжении всего эксперимента, в радиорезистентной раковой клеточной линии K562 и радиочувствительной раковой клеточной линии HCT116p; б – количество общих генов, чья экспрессия снижается на протяжении всего эксперимента, в радиорезистентной раковой клеточной линии K562 и радиочувствительной раковой клеточной линии HCT116p
Внутриклеточные механизмы, в которых функционируют отобранные гены в радиорезистентной (K562) и радиочувствительной (HCT116p) раковых клеточных линиях определяются с помощью онлайн программы PANTHER.

а б
Рис. 2. Общие сигнальные пути для радиорезистентной (K562)
и радиочувствительной (HCT116p) раковых клеточных линий:
а – общие сигнальные пути для двух раковых клеточных линий, в которые включены гены,
чья экспрессия увеличивалась на протяжении 1, 12 и 24 часов; б – общие сигнальные пути
для двух раковых клеточных линий, в которые включены гены,
чья экспрессия снижается на протяжении 1, 12 и 24 часов
На рис. 2 показано количество сигнальных путей, в которых задействованы отобранные гены и количество общих сигнальных путей для радиорезистентной (K562) и радиочувствительной (HCT116p) раковых клеточных линий. Для расчета активации сигнальных путей отобрано пять (EGF receptor signaling pathway, Apoptosis signaling pathway, FGF signaling pathway, Gonadotropin releasing hormone receptor pathway, Huntington disease), которые являются общими для двух раковых клеточных линий в случае, когда происходит повышение (рис. 2, а) и снижение (рис. 2, б) экспрессии генов на протяжении всего эксперимента.
В табл. 1, 2 приведены данные активации сигнальных путей в двух раковых клеточных линиях.
Таблица 1
Активация сигнальных путей в радиочувствительной раковой
клеточной линии HCT116p в течение 1, 12 и 24 часов после ионизирующего излучения
|
Сигнальные пути |
HCT 116p |
HCT 116p 12 часов |
HCT 116 p |
|
1. EGF receptor signaling pathway |
2,4361 |
–13,654 |
–5,8752 |
|
2. Apoptosis signaling pathway |
5,1936 |
3,2807 |
6,7716 |
|
3. FGF signaling pathway |
–6,5782 |
–11,7453 |
3,9147 |
|
4. Gonadotropin releasing hormone receptor pathway |
–12,2643 |
–21,4517 |
–9,1085 |
|
5. Huntington disease |
0,9516 |
8,2162 |
15,6322 |
Таблица 2
Активация сигнальных путей в радиорезистентной раковой клеточной линии K562 в течение 1, 12 и 24 часов после ионизирующего излучения
|
Сигнальные пути |
K562 1 час |
K562 12 часов |
K562 24 часа |
|
1. EGF receptor signaling pathway |
–2,902 |
–5,2527 |
6,7421 |
|
2. Apoptosis signaling pathway |
0,0887 |
3,1617 |
–1,8798 |
|
3. FGF signaling pathway |
–1,7543 |
1,5256 |
7,9806 |
|
4. Gonadotropin releasing hormone receptor pathway |
–3,6758 |
0,95 |
8,0356 |
|
5. Huntington disease |
–0,5998 |
3,2712 |
–1,8408 |
Активация сигнальных путей в радиорезистентной (K562) и радиочувствительной (HCT 116p) раковых клеточных линиях кардинально отличается. Сигнальные пути EGF receptor signaling pathway и Gonadotropin releasing hormone receptor pathway в радиочувствительной (HCT 116p) раковой клеточной линии имеют отрицательное значение активации (–5,8752; –9,1085) после 24 часов эксперимента, а в радиорезистентной (K562) раковой клеточной линии активация имеет положительное значение (6,7421; 8,0356). В Apoptosis signaling pathway и Huntington disease активация сигнальных путей имеет относительно высокие показатели в радиочувствительной раковой клеточной линии (6,7716; 15,6322) и минимальные значения в радиорезистентной раковой клеточной линии (–1,8798; –1,8408) после окончания эксперимента. Характер активации сигнального пути FGF signaling pathway в двух раковых клеточных линиях имеет схожую тенденцию к росту, но значения в радиорезистентной раковой клеточной линии выше (7,9806), чем в радиочувствительной раковой клеточной линии (3,9147). Эти данные доказывают, что при анализе транскриптома необходимо учитывать не только экспрессию отдельных генов, но и учитывать их работу в совокупности, т.к. изменение экспрессии одного гена способно привести к изменению функционирования всей системы генома.
Таким образом, найдены общие гены для радиорезистентной (K562) и радиочувствительной (HCT116p) раковых клеточных линий, чья экспрессия растет или падает на протяжении всего эксперимента. В результате последующего анализа обнаружены общие сигнальные пути, компонентами которых являются отобранные гены и отобрано 5 сигнальных путей, характерных для обеих клеточных линий. Количественный анализ активации сигнальных путей продемонстрировал, что, несмотря на многоступенчатый отбор по сходству адаптивного ответа радиорезистентной и радиочувствительной клеточной линии на радиационное воздействие, активация этих сигнальных путей имеет различную динамику и направленность.
Результаты получены в рамках программы «УМНИК» и в рамках выполнения государственного задания Минобрнауки РФ.
Рецензенты:Балыкин М.В., д.б.н., профессор, заведующий кафедрой адаптивной физической культуры Института медицины, экологии и физической культуры, ФГБОУ ВПО «Ульяновский государственный университет», г. Ульяновск;
Светухин В.В., д.ф.-м.н., профессор, научный руководитель научно-исследовательского технологического института УлГУ,
г. Ульяновск.
Работа поступила в редакцию 05.12.2014.
Библиографическая ссылка
Глущенко Е.С., Антонова А.В., Свеколкин В.П., Мякишева С.К., Федотова С.В., Слесарев С.М., Саенко Ю.В. КОЛИЧЕСТВЕННЫЙ АНАЛИЗ АКТИВАЦИИ СИГНАЛЬНЫХ ПУТЕЙ РАДИОРЕЗИСТЕНТНОЙ И РАДИОЧУВСТВИТЕЛЬНОЙ РАКОВЫХ КЛЕТОЧНЫХ ЛИНИЙ // Фундаментальные исследования. 2014. № 12-2. С. 307-311;URL: https://fundamental-research.ru/en/article/view?id=36222 (дата обращения: 24.06.2026).