


Scientific journal
Fundamental research
ISSN 1812-7339
"Перечень" ВАК
ИФ РИНЦ = 1,798
STUDY OF SPRING WHEAT REACTION TO SALINITY ON THE VARIABILITY OF SPROUTS MORPHOMETRIC PARAMETERS

Проблема солеустойчивости растений – одна из наиболее актуальных в растениеводстве. Необходимость ее всестороннего изучения определяется наличием во многих странах больших площадей засоленных почв, представляющих значительное препятствие для роста, развития и повышения урожайности сельскохозяйственных растений [2]. Площади засоленных земель имеют тенденцию к постоянному и существенному увеличению в результате процессов вторичного засоления, которое ежегодно приносит большой урон многим отраслям растениеводства и ограничивает расширение площадей под различные сельскохозяйственные культуры в засушливых районах нашей страны [8, 12, 15]. Согласно сведениям, приведенным И. Сабольчем в монографии «Моделирование процессов засоления и осолонцевания почв» [цит. по 11], общая площадь засоленных почв в мире составляет более 950 млн га.
Тюменская область характеризуется частой сменой разнотипных почв на ограниченном пространстве. Зональные черноземные почвы включают в себя солонцы луговые и дерновые, солончаки луговые, лугово-болотные [1]. Засоленные почвы составляют 1,6 % почвенного покрова и расположены в южной части области. Представлены они преимущественно лугово-черноземными солонцеватыми (25,3 % площади засоленных почв), луговыми солонцеватыми и солончаковатыми почвами (около 34 %), а также солонцами гидроморфными (23 %) и маршевыми засоленными почвами (около 11 %) [11].
Одним из путей снижения отрицательного воздействия на растения является подбор сортов, устойчивых к повышенному содержанию солей в почве. Для диагностики устойчивости растений к засолению почвы применяют целый ряд методов, и все они имеют в своей основе одно необходимое условие – создание провокационного фона засоления. Это связано с тем, что уровень солеустойчивости растений закреплен генетически и проявляется лишь при действии этого экстремального фактора [6].
При определении солеустойчивости используется ряд методик, различающихся способами проращивания семян, концентрациями растворов солей и элементами учета (скорость набухания семян в солевых растворах, энергия прорастания, всхожесть семян, длина и масса проростков и т.д.). Оценка устойчивости растений к засолению по прорастанию семян дает возможность сравнивать как близкородственные группы (сорта и виды одной культуры), так и биологически отдаленные (разные культуры) [10].
Анализ и обобщение результатов из литературных источников дает основание предположить, что изучение особенностей раннего онтогенеза с детальным анализом первичной корневой системы и надземных органов проростков по комплексу количественных признаков может представлять ценность для отбора высокоустойчивых к засолению форм культурных растений.
Цель нашего исследования – изучение особенностей роста и развития яровой мягкой пшеницы в условиях засоления с использованием различных методов диагностики.
Материалы и методы исследования
Экспериментальная часть работы выполнена на кафедре ботаники, биотехнологии и ландшафтной архитектуры Института биологии Тюменского государственного университета. Объектом изучения послужили 10 сортов яровой мягкой пшеницы (Triticum aestivum L.) (Авиада, Икар, Ильинская, Казахстанская 10, Латона, Лютесценс 70, Новосибирская 15, СКЭНТ 1, СКЭНТ 3, Тюменская 80), относящиеся к разновидностям lutescens и pirothrix.
Оценку сортов яровой мягкой пшеницы по солеустойчивости проводили с использованием двух методических подходов:
1) проращивание семян на солевых растворах в чашках Петри;
2) проращивание семян на засоленном песке в вегетационных сосудах.
Для создания хлоридного засоления использовали 1,40 %-й раствор NaCl, сульфатного – 2,78 %-й раствор Na2SO4, карбонатного – 2,06 %-й раствор Na2CO3, что соответствует осмотическому давлению растворов 1 МПа.
При первом подходе проращивание семян проводили в стерильных чашках Петри на фильтровальной бумаге, увлажненной дистиллированной водой (контроль) или растворами солей (опытные варианты). Объем выборки – 50 семян в трехкратной повторности для каждого варианта. Чашки с семенами помещали в термостат ТСО-1/80 СПУ с температурой 22–24 °С. На седьмой день эксперимента учитывали лабораторную всхожесть семян, длину побегов и корней, число корней, сырую и сухую массу побегов и корней.
При втором методическом подходе в качестве субстрата использовали прокаленный песок, увлажненный до 60–70 % полной влагоемкости, которым заполняли вегетационные сосуды из инертного материала. Засоление проводили при раскладке семян растворами солей. В контроле песок увлажняли дистиллированной водой. Объем выборки – 100 семян в трехкратной повторности для каждого варианта. Проращивание семян проводили в условиях искусственного освещения в универсальной камере для моделирования климатических условий среды
MLR-351 при температуре 24–26 °С и 50 %-й влажности. На десятый день эксперимента учитывали всхожесть семян, длину побегов и корней, число корней, длину и ширину первого листа, сырую и сухую массу побегов и зародышевых корней, рассчитывали площадь первого листа.
Расчет основных статистических параметров проводили по стандартным методикам [9], а также использовались программы для статистической обработки данных Statistica 6.0 и STATAN.
Результаты исследования
и их обсуждение
При оценке культурных растений стандартными лабораторными методами критериями солеустойчивости обычно являются такие показатели, как энергия прорастания, лабораторная всхожесть семян. Однако в научной литературе отмечается, что изменение показателей прорастания семян при засолении нередко слабо коррелирует со степенью солеустойчивости растений. В связи с этим для получения более достоверных и объективных результатов исследований рекомендуется кроме учета всхожести или энергии прорастания семян определять также количество зародышевых корней, длину проростков и корней, интенсивность роста проростков во времени [14].
Изучение адаптации растений к засолению показывает, что этот процесс идет неравномерно в течение онтогенеза [7]. Наибольшую чувствительность к высоким концентрациям солей растения проявляют на первых этапах развития, начиная с момента прорастания семян и появления всходов [3, 4].
По обобщенным результатам опытов Г.В. Удовенко и Э.А. Гончаровой [13], отрицательное действие повышенной концентрации солей сказывается уже на начальных этапах жизни растения. Проявляется это в задерживании набухания семян, при высоком уровне засоления (15–20 атм.) семена не наклевываются.
П.А. Генкель [5], сравнивая типы засоления (при низких уровнях 5–7 атм.), отмечал, что хлоридное засоление сильнее, чем сульфатное, угнетает рост и снижает продуктивность пшеницы, что связано с высокой токсичностью хлора для клеток. При более сильном засолении решающее значение имеет самоповышение осмотического давления, и различия исчезают.
В результате наших исследований установлено, что в условиях засоления в чашках Петри и вегетационных сосудах наблюдалось значительное снижение лабораторной всхожести семян: на фоне с NaCl на 25,3–37,9 %, на фоне с Na2SO4 на 47,7–58,4 % по сравнению с контролем (в среднем по сортам). В условиях карбонатного засоления в чашках Петри семена не проросли, в вегетационных сосудах отмечен 1 % проросших семян, что указывает на отрицательное воздействие фактора (рис. 1).
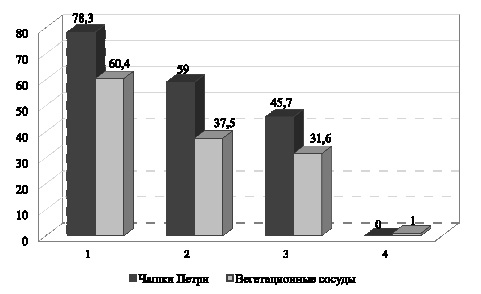
Рис. 1. Всхожесть семян пшеницы в разных вариантах эксперимента (среднее по 10 сортам), %. Примечание. Статистически достоверные различия
* – с контролем; ● – с NaCl при Р < 0,05; 1 – контроль; 2 – NaCl; 3 – Na2SO4; 4 – Na2CO3
Высокой лабораторной всхожестью семян в чашках Петри в контрольном варианте характеризовались сорта Авиада (92,7 %) и Казахстанская 10 (91,3 %); на фоне с NaCl – СКЭНТ 3 и Казахстанская 10, имевшие 80,7 % всхожих семян; на фоне с Na2SO4 выделились сорта Авиада и Тюменская 80 – 63,3 и 60,0 % соответственно. Наименьший процент взошедших семян отмечен у сорта СКЭНТ 1 (контроль – 48,0 %, фон с NaCl – 21,3 %, фон с Na2SO4 – 18,7 %).
В вегетационных сосудах высокие показатели лабораторной всхожести семян на фонах с хлоридным и карбонатным засолениями были отмечены у сортов Лютесценс 70 (NaCl – 71,3 %, Na2SO4 – 50,3 %) и Новосибирская 15 (NaCl – 51,0 %, Na2SO4 – 41,7 %). У сорта Лютесценс 70 по всхожести отмечены наименьшие отличия с контролем в условиях хлоридного засоления, составившие 12,7 %. Наибольшей чувствительностью к обоим типам засоления характеризовался сорт Авиада, у которого снижение процента всхожих семян по отношению к контролю составило 76,5–77,8 %.
Изменение линейных параметров ростовых процессов отражает солеустойчивость сортов более достоверно, чем оценка показателей прорастания семян, что связано с общей природой устойчивости растений. Подавление клеточного деления и особенно растяжения при действии неблагоприятных условий приводит к задержке роста растений, которую рассматривают как защитную реакцию. Устойчивые формы быстрее адаптируются к стрессу и восстанавливают рост [10]. Степень солеустойчивости определяется величиной отклонения выбранных параметров от контроля под влиянием засоления.
В нашем эксперименте засоление привело к угнетению большинства морфометрических параметров, особенно у проростков в чашках Петри (табл. 1).
По числу корней в среднем по сортам, при сравнении с контролем, в чашках Петри отмечено снижение признака в опытных вариантах на 24,5–36,7 %, в то время как в вегетационных сосудах – превышение на 4,2–6,3 %. Сорт СКЭНТ 3 во всех вариантах обоих экспериментов формировал наибольшее число корней. При сравнении двух типов засоления установлено, что сульфатное засоление в чашках Петри приводило к значительному снижению числа корней, в вегетационных сосудах различий не выявлено.
Таблица 1
Влияние различных типов засоления на проявление
морфометрических параметров проростков пшеницы
|
Признаки |
Контроль |
NaCl |
Na2SO4 |
||
|
Х ± mx |
Х ± mx |
±, % |
Х ± mx |
±, % |
|
|
Чашки Петри (в среднем по 7 сортам) |
|||||
|
Число корней, шт. |
4,9 ± 0,07 |
3,7 ± 0,07* |
–24,5 |
3,1 ± 0,06*● |
–36,7 |
|
Длина корней, мм |
116,9 ± 3,15 |
23,5 ± 0,77* |
–79,9 |
7,7 ± 0,35*● |
–93,4 |
|
Длина побегов, мм |
84,2 ± 1,94 |
4,8 ± 0,36* |
–94,3 |
2,2 ± 0,05*● |
–97,4 |
|
Вегетационные сосуды (в среднем по 10 сортам) |
|||||
|
Число корней, шт. |
4,8 ± 0,04 |
5,1 ± 0,04* |
+6,3 |
5,0 ± 0,05 |
+4,2 |
|
Длина корней, мм |
116,3 ± 2,45 |
45,8 ± 0,68* |
–60,6 |
42,4 ± 0,63*● |
–63,5 |
|
Длина побегов, мм |
168,0 ± 2,72 |
69,8 ± 1,82* |
–58,5 |
65,5 ± 1,81* |
–61,0 |
|
Длина листа, мм |
133,9 ± 2,45 |
41,5 ± 1,70* |
–69,0 |
37,5 ± 1,62* |
–72,0 |
|
Ширина листа, мм |
2,9 ± 0,03 |
2,5 ± 0,02* |
–13,8 |
2,4 ± 0,03* |
–17,2 |
|
Площадь листа, мм2 |
335,3 ± 7,24 |
89,5 ± 3,84* |
–73,3 |
81,2 ± 3,61* |
–75,8 |
Примечание. Статистически достоверные различия * – с контролем, • – с NaCl при Р<0,05; ± , % – отклонение от контроля.
При оценке сортов в чашках Петри на фоне без засоления длина первичных корней варьировала от 88,7 ± 3,53 мм (СКЭНТ 3) до 150,7 ± 9,12 мм (Латона), при среднем значении по сортам 116,9 ± 3,15 мм. При проращивании семян на солевых растворах длина зародышевых корней уменьшалась до 7,7 ± 0,35 мм (в варианте с Na2SO4) – 23,5 ± 0,77 мм (в варианте с NaCl). Наименьшее отклонение от контроля (на 39,0– 76,7 %) по данному показателю в вариантах с засолением отмечено у сортов Казахстанская 10 и Латона. Значительное угнетение корней по данному признаку на фоне с NaCl обнаружено у сорта Тюменская 80, на фоне с Na2SO4 – у сорта СКЭНТ 1.
В вегетационном эксперименте длина первичных корней в контроле изменялась от 32,0 ± 3,33 мм (СКЭНТ 1) до 153,4 ± 4,00 мм (Икар) (среднее по сортам 116,3 ± 2,45 мм). В опытных вариантах по данному показателю отличия между действием различных типов засоления были менее выражены по сравнению с экспериментом в чашках Петри. У проростков на фоне с NaCl первичная корневая система в стрессовых условиях по длине уменьшилась на 60,6 %, на фоне с сульфатным засолением – на 63,5 %.
При проращивании семян в чашках Петри проростки формировали более короткие побеги (84,3 ± 1,94 мм) по сравнению с побегами в вегетационных сосудах (168,0 ± 2,72 мм). Сульфатное засоление в чашках Петри приводило к значительному уменьшению побегов по сравнению с хлоридным, длина которых в среднем по сортам составила соответственно 2,2 ± 0,05 и 4,8 ± 0,36 мм. На прокаленном песке данный показатель в опытных вариантах изменялся от 65,5 ± 1,81 мм (на фоне с Na2SO4) до 69,8 ± 1,82 мм (на фоне с NaCl) (табл. 1).
Вегетационный эксперимент позволил нам сравнить влияние различных типов засоления на морфометрические параметры первого настоящего листа у разных сортов, что невозможно при проведении эксперимента в чашках Петри. В контрольном варианте минимальная площадь листа отмечена у сорта СКЭНТ 1 (77,03 мм2), имевшего короткую и узкую листовую пластинку. Сорт Новосибирская 15 превосходил другие сорта по длине, ширине и площади листовой пластинки (466,73 мм2).
Проростки опытных вариантов имели короткие и узкие листовые пластинки. По площади листовой пластинки в варианте с хлоридом натрия выделился сорт Казахстанская 10 (167,84 мм2), имевший наибольшую длину листа. Минимальная длина, ширина и площадь первого настоящего листа (21,74 мм2) отмечена у сорта Авиада.
При изучении морфометрических параметров проростков в условиях сульфатного засоления установлено, что сорт Казахстанская 10 характеризовался более развитыми побегами (длина побега – 102,0 мм, длина листа – 68,4 мм, площадь листа – 131,36 мм2). Значительное угнетение надземной сферы характерно для сортов СКЭНТ 1 и Авиада (длина побега – 31,6–41,6 мм, длина листа – 12,4–19,1 мм, ширина листа – 1,5–2,1 мм, площадь листовой пластинки – 21,74–23,92 мм2). По фенотипическому проявлению всех морфологических признаков опытные проростки уступали контрольным.
В условиях засоления отмечено увеличение вариабельности большинства изученных параметров проростков, особенно при оценке солеустойчивости сортов пшеницы в чашках Петри (табл. 2). Возможно, это связано с тем, что при данном способе оценки и корневая система, и побеги контактировали с ионами солей, тогда как в вегетационных сосудах –
только корни.
Таблица 2
Изменчивость морфометрических параметров проростков пшеницы под влиянием различных типов засоления (CV, %)
|
Признаки |
Контроль |
NaCl |
Na2SO4 |
|
Чашки Петри (в среднем по 7 сортам) |
|||
|
Число корней, шт. |
19,78 |
27,74 |
29,90 |
|
Длина корней, мм |
38,98 |
47,20 |
65,26 |
|
Длина побегов, мм |
33,99 |
109,56 |
36,52 |
|
Вегетационные сосуды (в среднем по 10 сортам) |
|||
|
Число корней, шт. |
15,58 |
14,62 |
14,92 |
|
Длина корней, мм |
36,19 |
25,08 |
25,08 |
|
Длина побегов, мм |
27,80 |
44,03 |
46,69 |
|
Длина листа, мм |
36,36 |
69,25 |
72,55 |
|
Ширина листа, мм |
15,91 |
15,86 |
20,65 |
|
Площадь листа, мм2 |
36,97 |
72,50 |
74,18 |
В меньшей степени в условиях засоления варьировали число корней, независимо от способа оценки, и ширина листа в вегетационном эксперименте. На фоне с хлоридным засолением в чашках Петри наибольшей изменчивостью характеризовалась длина побегов, в сосудах на прокаленном песке – длина и площадь листовой пластинки. В условиях сульфатного засоления при проращивании семян на фильтровальной бумаге сильнее варьировала длина корней, на песке – длина и площадь первого настоящего листа. Коэффициент вариации на фоне с Na2SO4 изменялся от 14,92 до 74,18 %, на фоне с NaCl
от 14,62 до 72,50 %.
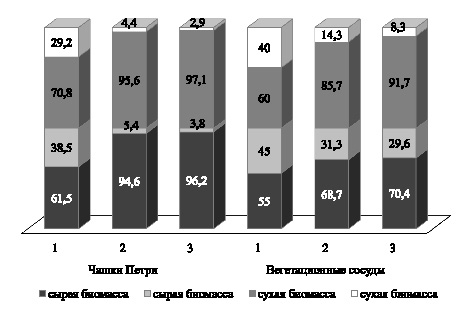
Рис. 2. Соотношение корневой системы и побегов в сырой и сухой биомассе проростков, %. Примечание: 1 – контроль; 2 – NaCl; 3 – Na2SO4
Анализ сырой и сухой биомассы показал, что в стрессовых условиях происходит снижение сырой массы зародышевых корней, сырой и сухой массы побегов, независимо от типов засоления и использованной методики оценки. Напротив, сухая масса корней в ряде случаев увеличивается, что мы связываем с повреждением корневой системы и накоплением ионов солей.
В среднем по сортам в условиях засоления в структуре биомассы сырых и сухих проростков преобладали корни как в чашках Петри, так и в вегетационных сосудах. При выращивании на песке проростки имели более развитые побеги и соответственно высокую сырую и сухую массу по сравнению с растениями, полученными при проращивании семян на солевых растворах (рис. 2).
С целью выявления солеустойчивых сортов нами проведено балловое ранжирование, основанное на учете процента снижения изученных признаков по отношению к контролю. По результатам исследования на фоне с NaCl в чашках Петри выделены сорта СКЭНТ 3, Казахстанская 10, Латона, Лютесценс 70, на фоне с Na2SO4 – Икар, Казахстанская 10, СКЭНТ 3, Тюменская 80 и Лютесценс 70, угнетавшиеся в меньшей степени по сравнению с контролем по большинству изученных признаков. Наиболее чувствительным к хлоридному и сульфатному засолению оказался
сорт СКЭНТ 1.
В вегетационных сосудах к числу наиболее устойчивых на хлоридном засолении отнесены сорта Казахстанская 10, СКЭНТ 1, Тюменская 80, на сульфатном – Казахстанская 10, Латона, Лютесценс 70. Наибольшую чувствительность к действию NaCl и Na2SO4 проявил сорт Авиада.
Таким образом, при выявленной общей закономерности угнетения ростовых процессов проростков яровой пшеницы под воздействием солей сорта значительно различались между собой по проявлению ряда признаков. Максимальной солеустойчивостью обладал сорт Казахстанская 10 независимо от типа засоления и способа оценки. Кроме того, по результатам баллового ранжирования по комплексу показателей выделены сорта с минимальным снижением признаков в условиях стресса – Лютесценс 70, СКЭНТ 3, Тюменская 80. К числу сортов с низкими результатами в первом эксперименте отнесен СКЭНТ 1, во втором – Авиада.
Заключение
В результате проведения оценки сортов яровой пшеницы по комплексу признаков установлено, что все типы засоления оказывают отрицательное воздействие, вызывая снижение морфометрических параметров проростков. При этом карбонатное засоление является самым губительным, а сульфатное действует сильнее, чем хлоридное, на первых этапах онтогенеза
растений.
При использовании двух методов диагностики не получено одинаковых результатов в отношении солеустойчивости изученных сортов яровой пшеницы. Постановка эксперимента в чашках Петри отличается меньшей продолжительностью и трудоемкостью, что позволяет провести оценку большого объема материала. Выращивание растений в вегетационных сосудах дает возможность учета большего количества параметров, что, на наш взгляд, повышает точность
эксперимента.
Белкина Р.И., д.с.-х.н., профессор кафедры технологии производства, хранения и переработки продукции растениеводства, ФГБОУ ВПО «Государственный аграрный университет Северного Зауралья», г. Тюмень;
Турсумбекова Г.Ш., д.с.-х.н., профессор кафедры общей биологии, ФГБОУ ВПО «Государственный аграрный университет Северного Зауралья», г. Тюмень.
Работа поступила в редакцию 05.12.2014.
Библиографическая ссылка
Белозерова А.А., Боме Н.А. ИЗУЧЕНИЕ РЕАКЦИИ ЯРОВОЙ ПШЕНИЦЫ НА ЗАСОЛЕНИЕ ПО ИЗМЕНЧИВОСТИ МОРФОМЕТРИЧЕСКИХ ПАРАМЕТРОВ ПРОРОСТКОВ // Фундаментальные исследования. 2014. № 12-2. С. 300-306;URL: https://fundamental-research.ru/en/article/view?id=36221 (дата обращения: 24.06.2026).