


Scientific journal
Fundamental research
ISSN 1812-7339
"Перечень" ВАК
ИФ РИНЦ = 1,798
DEPENDENCE OF PHOTOSYNTHETIC RESPONSE ON VARIATION POTENTIAL AMPLITUDE ON WHEAT SEEDLING

Электрогенез, т.е. способность к генерации электрических потенциалов, является универсальным свойством живых организмов, включая растения [2, 3]. При локальном действии неблагоприятных факторов у растений возникают электрические сигналы, проходящие в интактные участки растения [2]. В настоящее время чаще всего выделяют два типа электрических сигналов у высших растений [2, 3]: возникающий при действии неповреждающих стимулов потенциал действия (ПД) и индуцируемый повреждающими воздействиями вариабельный потенциал (ВП).
ПД представляет собой импульсную реакцию, возникающую в ответ на действие умеренных стимулов и способную к активному распространению по проводящим пучкам растения [2]. Основной механизм ПД связан с потоками кальция, хлора и калия, однако участие протонной АТФазы в процессе представляется также возможным [1]. ВП является электрической реакцией вариабельной формы и связан прежде всего с обратимой инактивацией протонной АТФазы [2]. Распространение ВП связано с распространением по проводящим пучкам неэлектрического сигнала гидравлической, химической или комплексной природы [6, 10].
С другой стороны, известно, что при локальном действии неповреждающих и повреждающих стимулов в интактных участках растения могут развиваться разнообразные функциональные ответы, включая изменения фотосинтетической активности [7], т.е. существуют быстрые стрессовые сигналы, распространяющиеся по растительному организму. Электрические сигналы являются наиболее вероятным кандидатом на роль таких сигналов [3], однако экспериментальные доказательства такого участия ограничены. Так, в случае ВП было показано, что инактивация фотосинтеза развивается только при прохождении сигнала в лист [8], а времена начала развития фотосинтетического ответа в разных частях растения отражают динамику распространения вариабельного потенциала [5]. При этом совершенно неисследованной остается зависимость фотосинтетического ответа от параметров электрического сигнала, в первую очередь его амплитуды. Таким образом, целью настоящей работы стало исследование зависимости параметров фотосинтетического ответа от амплитуды вариабельного потенциала у проростков пшеницы.
Материалы и методы исследования
В работе использовались проростки мягкой пшеницы (Triticum aestivum L.) возрастом 14–15 суток, выращенные гидропонным способом на 50 % среде Хогланда – Арнона в климатической камере KBW-240 («Binder») при температуре 24 °С и 16-часовом световом периоде.
ВП вызывали ожогом открытым пламенем кончика второго листа пшеницы в течение 2 с. Исследования фотосинтетической и электрической активности проводились на втором листе.
Электрофизиологические измерения проводились методом экстраклеточной регистрации с использованием двух пар неполяризующихся хлорсеребряных макроэлектродов ЭВЛ-1M3, высокоомного милливольтметра ИПЛ-113 и ПК. Первый (Э1) и второй (Э2) измерительные электроды располагались на расстоянии 4 и 8 см от места ожога соответственно. Электрод сравнения находился в жидкости, контактировавшей с корнями растения.
Параметры фотосинтеза, включая скорость ассимиляции CO2 (A), нефотохимическое тушение флуоресценции (NPQ) и квантовые выходы фотосистем I (γ(PSI)) и II (γ(PSII)), были определены с использованием стандартной системы для фотосинтетических исследований, включающей в себя газоанализатор GFS-3000, РАМ-флуориметра Dual-PAM-100 и измерительного блока Cuvette 3010-Dual. Измерение проводилось на участке листа, расположенном между электродами. Темновая адаптация перед включением света и время освещения перед ожогом составляли по 30 мин каждое.
Эксперименты проводились в восемнадцатикратной биологической повторности. При анализе связи параметров фотосинтеза и амплитуды ВП все 18 результатов ранжировались по возрастанию амплитуды сигнала под первым или вторым электродом. После этого каждый набор данных делился на три равные группы (по 6 экспериментов для низкой, средней и высокой амплитуды ВП), для каждой из которых оценивались средние значения электрических и фотосинтетических параметров.
Результаты исследования
и их обсуждение
На первом этапе исследования была изучена способность локальных ожогов вызывать распространение электрических сигналов у проростков пшеницы и вызывать у них фотосинтетические ответы. Локальные ожоги вызывали у проростков пшеницы распространение электрических сигналов сложной формы, включающей в себя как быструю деполяризацию, так и более медленные изменения (рис. 1). Амплитуда реакции варьировала в широком пределе и составляла от 10 до 90 мВ, при этом величина амплитуды снижалась по мере удаления от зоны ожога. Скорость распространения сигнала составляла около 2 мм∙с–1 и также характеризовалась высокой вариабельностью (0,5–3 мм∙с–1). Такие признаки – возникновение при действии ожога, сложная форма, уменьшение амплитуды по мере удаления от зоны повреждения, относительно низкая скорость распространения и высокая вариабельность параметров – позволяют идентифицировать обнаруженный сигнал как ВП [2, 10].
Через две-три минуты после распространения ВП в листе начинал развиваться фотосинтетический ответ, который представлял собой инактивацию фотосинтеза (рис. 1). При этом происходило снижение скорости ассимиляции углекислого газа (на 1–3 мкмоль∙м–2∙с–1), рост нефотохимического тушения флуоресценции (на 0,2–0,8 единиц) и небольшое уменьшение квантовых выходов обеих фотосистем. Развитие подобного ответа очень хорошо согласуется с широким спектром работ, в которых показано, что локальные повреждения существенно снижают фотосинтетическую активность у растений различных видов активности [3, 5, 7, 8], что, по-видимому, опосредуется распространением электрических сигналов [5, 8]. Тот факт, что в наших экспериментах развитие фотосинтетического ответа происходило после распространения ВП, свидетельствует в пользу того, что именно распространение вариабельного потенциала может подавлять фотосинтетическую активность.
Однако не может быть полностью исключен и вариант независимого распространения ВП и какого-либо иного сигнала, влияющего на фотосинтетическую активность. Такая возможность была показана, в частности, для кукурузы [4], исследования которой показали, что на фотосинтез помимо электрического сигнала может влиять также гидравлическая волна, вызванная действием повреждающего стимула. В то же время, если распространение ВП и развитие фотосинтетического ответа являются независимыми процессами, то можно ожидать, что зависимость параметров снижения фотосинтеза от параметров вариабельного потенциала будет выражена слабо.
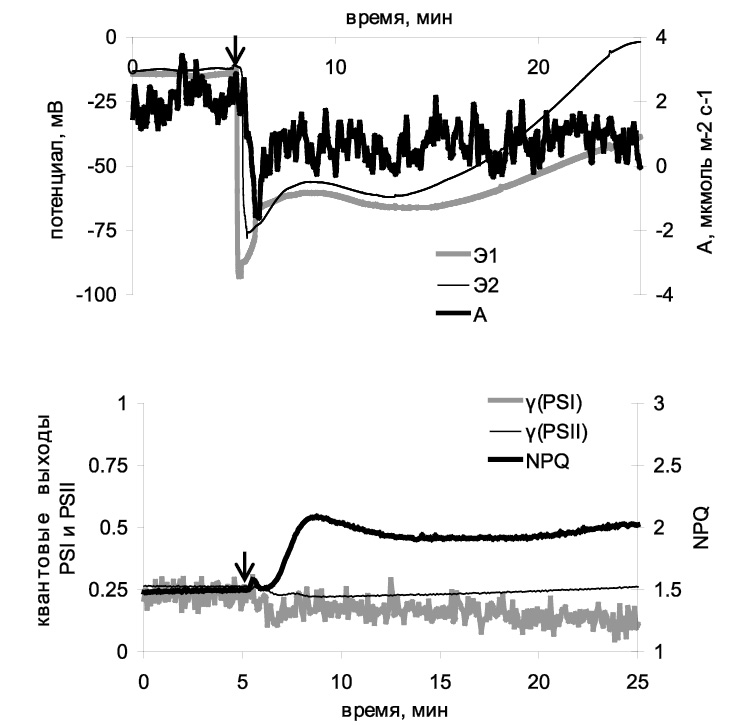
Рис. 1. Типичный пример изменений электрической и фотосинтетической активности, вызванных ожогом кончика листа пшеницы. Стрелкой обозначен момент нанесения ожога
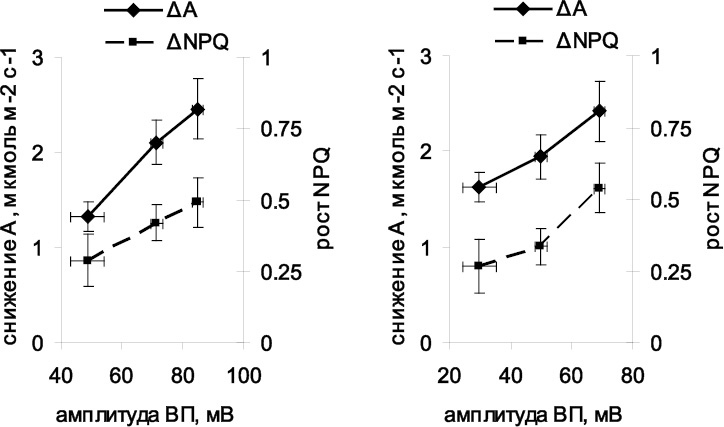
а б
Рис. 2. Зависимости снижения ассимиляции и возрастания нефотохимического тушения флуоресценции от амплитуды вариабельного потенциала,
измеренной первым (а) и вторым (б) электродами
На следующем этапе работы была проанализирована зависимость уменьшения скорости ассимиляции углекислого газа и возрастания нефотохимического тушения флуоресценции от амплитуд ВП под первым и вторым электродами (рис. 2). Показано, что как величина снижения ассимиляции, так и величина возрастания NPQ повышаются при увеличении амплитуд ВП под первым и вторым электродами. Форма обнаруженных зависимостей близка к линейной, что говорит о существовании прямой пропорциональности между величиной ВП и величиной фотосинтетического ответа. Полученные результаты хорошо согласуются с полученными нами ранее данными о том, что между амплитудой ВП и связанными с ней изменениями вне- и внутриклеточной рН имеется высокая положительная корреляция [9], а также с гипотезой о ключевой роли входа протонов в развитии фотосинтетического ответа при распространении электрических сигналов у высших растений [5, 9].
Заключение
Полученные нами результаты показывают, что локальные повреждения вызывают у проростков пшеницы генерацию ВП, которые распространяются по растению. В свою очередь при распространении ВП вызывают инактивацию фотосинтетических процессов, которая проявляется в уменьшении скорости ассимиляции углекислого газа, росте нефотохимического тушения и небольшом снижении квантовых выходов фотосистем. Между величиной фотосинтетического ответа и амплитудой ВП существует положительная связь: при небольшой амплитуде ВП наблюдаются относительно маленькие изменения фотосинтеза и наоборот. Форма зависимости параметров фотосинтетического ответа от амплитуды ВП вблизи зоны регистрации фотосинтеза близка к линейной.
В целом наши результаты подтверждают, что именно распространение ВП является механизмом, с помощью которого происходит развитие ответа в неповрежденных участках растения при действии на него локальных стимулов.
Работа выполнена при финансовой поддержке Российского научного фонда (проект № 14-26-00098).
Рецензенты:Охапкин А.Г., д.б.н., профессор, зав. кафедрой ботаники биологического факультета, ФГАОУ ВПО «Нижегородский государственный университет им. Н.И. Лобачевского» Министерства образования и науки РФ, г. Нижний Новгород;
Бережная Г.А., д.б.н., проф., профессор кафедры ботаники, физиологии и защиты растений, ФГБОУ ВПО «Нижегородская государственная сельскохозяйственная академия» Министерства сельского хозяйства РФ, г. Нижний Новгород.
Работа поступила в редакцию 28.11.2014.
Библиографическая ссылка
Шерстнева О.Н, Сурова Л.М, Воденеев В.А, Сухов В.С ЗАВИСИМОСТЬ ФОТОСИНТЕТИЧЕСКОГО ОТВЕТА ОТ АМПЛИТУДЫ ВАРИАБЕЛЬНОГО ПОТЕНЦИАЛА У ПРОРОСТКОВ ПШЕНИЦЫ // Фундаментальные исследования. 2014. № 11-12. С. 2661-2664;URL: https://fundamental-research.ru/en/article/view?id=36041 (дата обращения: 17.06.2026).