


Scientific journal
Fundamental research
ISSN 1812-7339
"Перечень" ВАК
ИФ РИНЦ = 1,798
MORPHOLOGICAL FEATURES OF PAROTID SALIVARY GLAND AND ENZYMATIC ACTIVITIES ZONES STRUCTURES OF NEUROMUSCULAR SYNAPSES LATERAL MASSETER OF WHITE RATS ON DISPERSED FOOD FEEDING

Жевательная нагрузка является важнейшим механическим фактором морфогенеза органов ротовой полости. Она способствует формированию и адаптивной перестройке целого комплекса взаимодействующих органов и тканей полости рта и челюстно-лицевой области, совместно обеспечивающих выполнение важнейшей функции жизнеобеспечения – жевания [1, 2]. Изменение характера жевательной нагрузки вследствие изменения физических свойств пищи запускает механизмы гистоморфологической перестройки органов ротовой полости, адаптируя их к новым условиям функционирования. Исследования последних лет [4, 5, 6, 7] свидетельствуют о существенном влиянии диспергированной пищи не только на гистологическую структуру и постнатальный морфогенез отделов пищеварительного канала, но и органов, функционально связанных с ним и расположенных за пределами его стенки.
Целью исследования явилось изучение влияния воздействия диспергированой пищи на гистоморфологические особенности околоушной слюнной железы и структуру ферментоактивных зон нервно-мышечных синапсов латеральной жевательной мышцы 240-суточных белых крыс.
Материал и методы исследования
Материалом исследований послужили 16 самцов белых неинбредных крыс. На 21-е сутки постнатального развития животных произвольно распределяли на две группы – контрольную и опытную. Животных контрольной группы содержали в обычных условиях вивария на естественном для грызунов корме (зерно, овощи), для животных опытной группы аналогичную пищу подвергали механической обработке до мягкой пастообразной консистенции. Непосредственными объектами исследования были избраны латеральная жевательная мышца, сочетающая протракцию нижней челюсти с аддукцией и околоушная слюнная железа. Латеральная жевательная мышца и околоушная железа взяты у животных обеих экспериментальных групп на 240-е сутки постнатального онтогенеза.
Для изучения активности ацетилхолинэстеразы (АХЭ) в области нервно-мышечного синапса (НМС) использовали гистоэнзимохимическую методику выявления данного фермента тиоуксусной кислотой в модификации Г.М. Николаева и В.В. Шилкина [3]. При качественной характеристике АХЭ-позитивных зон оценивали выраженность и топографию конечного продукта реакции. Количественный анализ проводили на стандартной площади среза (87 тыс. мкм2) с помощью компьютерной программы специальной морфометрии биологических структур «Мекос–Ц1». При этом определяли: общее количество выявляемых АХЭ-позитивных зон; долю простых и сложных АХЭ-позитивных зон; количество и диаметр мышечных волокон; среднюю площадь сечения области нейромышечных синапсов для различных конструкций АХЭ-позитивных зон; форм-фактор площади сечения нейромышечных синапсов, позволяющий оценивать форму выявляемых сечений как веретенообразную (значение показателя ≤ 0,45), овоидную (0,46–0,64) или округлую (≥ 0,65).
Описание, сравнительно-морфологический анализ и морфометрия структур околоушных слюнных желез производились на гистологических препаратах, окрашенных гематоксилин-эозином Майера. Для морфометрического исследования использовалась компьютерная система, включающая микроскоп, цифровую видеокамеру JVC (Victor company, Япония) и компьютерную программу денситофотометрии «Мекос-Ц1» (Россия). Также определялись вес животных, индивидуальное количество потребляемой пищи в граммах, время, затрачиваемое на прием пищи в минутах, и количество подходов к еде. Полученные морфометрические данные подвергали статистической обработке с помощью пакета прикладной компьютерной программы «Statistica 6.0» (StatSoft Inc., USA).
Результаты исследований и их обсуждение
Результаты морфометрии околоушной слюнной железы свидетельствуют о том, что 240-е сутки постнатального онтогенеза характеризуются незначительными различиями (р > 0,05) в показателях площади сечения ацинусов животных, питающихся диспергированной пищей, и контрольных животных. Это, возможно, связано с потреблением большего количества пищи животными опытной группы по сравнению с контрольными. В частности, в ходе исследования установлено, что к концу исследуемого периода онтогенеза (240-е сутки) животные опытной группы потребляли пищи в 2,3 раза больше (p < 0,001), чем животные контрольной группы. Такое «переедание» животных опытной группы мы склонны связывать с более поздним наступлением преабсорбтивного (сенсорного) насыщения вследствие более позднего наступления рефлекторных влияний от механорецепторов слизистой оболочки полости рта, обусловленных слабым раздражающим действием диспергированной пищи на механорецепторы. Это подтверждается увеличением в 1,5 раза времени приема пищи животными опытной группы по отношению к контрольным животным и отсутствием различий в количестве подходов к еде контрольных и опытных животных. Примечательно, что вес 240-суточных контрольных животных составлял 532,90 ± 15,91 г, а животные опытной группы весили 453,57 ± 8,31 г (p < 0,05). Показано, что частота приемов пищи имеет минимальное влияние на накопление жировой клетчатки тела, но физические свойства пищи являются причиной увеличения ее эффективности и усвояемости и играют важную роль в процессе накопления жира [11]. Показано, что существует положительная корреляция между количеством стимулированной слюны, которая вырабатывается в процессе жевания, и увеличением размеров клеток околоушной слюнной железы и ее веса, а также индекса массы тела [10, 12].
В доступной литературе имеются сведения, что содержание 180-суточных крыс в течение 2 недель на жидкой диете не влияет на морфологические особенности вставочных и исчерченных протоков [13]. Результаты проведенного нами исследования свидетельствуют о том, что значения площади сечения стенок вставочных и исчерченных протоков 240-суточных животных, питающихся диспергированной пищей, значительно уступают таковым контрольных животных, что обусловлено более низкими значениями площади сечения цитоплазмы эпителиоцитов. В частности, значения площади сечения стенок вставочных протоков 240-суточных контрольных и опытных животных составляют 376,40 ± 7,87 мкм2 и 311,45 ± 7,04 мкм2 соответственно (p < 0,01). Площадь сечения стенок исчерченных протоков 240-суточных животных контрольной группы достигает 523,96 ± 9,61 мкм2, тогда как соответственный показатель животных опытной группы равен 469,14 ± 8,63 мкм2 (p < 0,01).
Латеральная жевательная мышца (m.masseter lateralis) тянется от предскуловой площадки верхнечелюстной кости от переднего отрезка скуловой дуги вниз и назад, оканчиваясь на линии по передневентральному краю массетерной площадки нижней челюсти (под прикрытием поверхностной порции) и сочетает протракцию нижней челюсти с аддукцией.
Латеральная жевательная мышца 240-суточных белых крыс как контрольной, так и опытной группы характеризуется высокой активностью ацетилхолинэстеразы (АХЭ) в области нервно-мышечных синапсов. Об этом свидетельствует 20-минутная инкубация срезов в рабочем растворе, обусловившая в ходе исследования появления конечного продукта реакции с мелкодисперсной структурой, четкими границами и насыщенной коричневой окраской. Для мышцы характерно наличие внесинаптической активности АХЭ.
Установлено, что в изучаемой мышце как опытной, так и контрольной групп ферментоактивные зоны (ФАЗ) нервно-мышечных синапсов расположены полями. Какая-либо закономерность локализации полей в пределах мышечного брюшка не выявлена (рис. 1).
Анализ особенностей морфологии АХЭ-позитивных зон нейро-мышечных синапсов мышцы позволяет выделить два основных типа конструкций ФАЗ: простые, характеризующиеся гомогенной локализацией активного фермента, и сложные, отличающиеся трабекулярной локализацией АХЭ с замкнутым или незамкнутым контуром, ограничивавшим ферментонегативную зону (рис. 2).
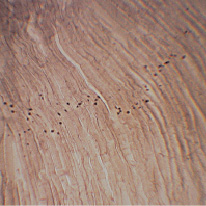
Рис. 1. НМС латеральной жевательной мышцы 240-суточных крыс в условиях измененной функциональной нагрузки. Окраска тиоуксусной кислотой, ×50
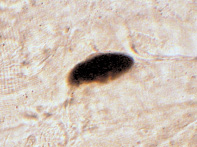
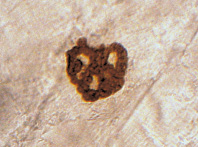
Рис. 2. НМС с простой АХЭ-позитивной зоной (слева) и со сложной АХЭ-позитивной зоной (справа) в латеральной жевательной мышце 240-суточных крыс в условиях измененной функциональной нагрузки. Окраска тиоуксусной кислотой, ×600
По данным наших исследований при питании диспергированной пищей в латеральной жевательной мышце 240-суточных животных отмечается преобладание относительного содержания НМС со сложной конструкцией ФАЗ, в то время как исследуемый мускул контрольных животных отличается большим относительным содержанием НМС с простой конструкцией ФАЗ (рис. 3). Преобразование простых ФАЗ НМС в сложные, по мнению Филимонова (1996), свидетельствует о повышении степени дифференцированности иннервационного аппарата мышцы [8]. Возможно, у опытных животных латеральная жевательная мышца совершает более сложные дифференцированные движения, связанные с обработкой в ротовой полости диспергированной пищи, несвойственной для данного типа животных.
Наряду с этим у животных опытной группы отмечается более высокий показатель средней площади сечения простых и сложных ФАЗ по сравнению с таковым контрольных животных. Увеличение площади контакта в нервно-мышечном синапсе свидетельствует о повышении концентрации химически активных веществ, выделяемых в этом участке, и, как следствие, о более энергичном мышечном сокращении [9].
В латеральной жевательной мышце число ФАЗ нервно-мышечных синапсов, приходящихся на одно мышечное волокно (∑ФАЗ/∑МВ), а также совокупная площадь ФАЗ в расчете на одно мышечное волокно (∑Sфаз/∑МВ) у животных опытной группы превышает данный показатель животных контрольной группы (р ≤ 0,05) (таблица). Более плотное расположение нервно-мышечных синапсов в мускуле, высокий показатель отношения площади нервной бляшки к поперечнику мышечного волокна отмечается в скелетной мускулатуре в условиях повышенной функциональной нагрузки [8].
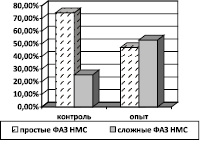
Рис. 3. Относительное содержание простых и сложных ФАЗ НМС латеральной жевательной мышцы 240-суточных белых крыс при питании диспергированной пищей (опыт) и в норме (контроль)
Интегративные показатели системы «двигательное окончание – мышечное волокно» латеральной жевательной мышцы 240-суточных животных в норме (контроль) и в условиях измененной функциональной нагрузки (опыт)
|
Показатели |
Среднее число ФАЗ, приходящихся на одно мышечное волокно (∑ФАЗ/∑МВ) |
Совокупная площадь ФАЗ в расчете на одно мышечное волокно (∑SФАЗ/∑МВ), мкм² |
|
Опыт |
1,08 ± 0,02* |
319,84 ± 15,38* |
|
Контроль |
0,73 ± 0,01 |
196,09 ± 9,64 |
Примечание. * – статистически значимое отличие показателей опытной группы от контрольных значений (р ≤ 0,05).
Заключение
Длительное питание диспергированной пищей вызывает уменьшение площади сечения эпителиоцитов вставочных и исчерченных протоков, что влечет значительное уменьшение площади сечения стенок указанных выводных протоков околоушной слюнной железы белых крыс. Питание диспергированной пищей обуславливает изменение функциональной роли латеральной жевательной мышцы в ходе жевания, проявляющееся в более сложных дифференцированных движениях данного мускула, связанных с обработкой в ротовой полости диспергированной пищи, несвойственной для данного типа животных. Следствием адаптации латеральной жевательной мышцы к изменению функциональной нагрузки явилось увеличение относительного количества сложных ФАЗ нервно-мышечных синапсов и усложнение их конструкции, возрастанием относительного числа и средней площади ФАЗ, приходящихся на одно мышечное волокно.
Рецензенты:
Балыкин М.В., д.б.н., профессор, заведующий кафедрой адаптивной физической культуры Института медицины, экологии и физической культуры, ФГБОУ ВПО «Ульяновский государственный университет», г. Ульяновск;
Слесарева Е.В., д.м.н., доцент, заведующая кафедрой морфологии Института медицины, экологии и физической культуры, ФГБОУ ВПО «Ульяновский государственный университет», г. Ульяновск.
Работа поступила в редакцию 15.07.2014.
Библиографическая ссылка
Курносова Н.А., Семенова М.А., Дрождина Е.П., Гальчин А.В., Чернова А.О. МОРФОЛОГИЧЕСКИЕ ОСОБЕННОСТИ ОКОЛОУШНОЙ СЛЮННОЙ ЖЕЛЕЗЫ И СТРУКТУР ФЕРМЕНТОАКТИВНЫХ ЗОН НЕРВНО-МЫШЕЧНЫХ СИНАПСОВ ЛАТЕРАЛЬНОЙ ЖЕВАТЕЛЬНОЙ МЫШЦЫ БЕЛЫХ КРЫС ПРИ ПИТАНИИ ДИСПЕРГИРОВАННОЙ ПИЩЕЙ // Фундаментальные исследования. 2014. № 9-5. С. 1027-1031;URL: https://fundamental-research.ru/en/article/view?id=35010 (дата обращения: 31.07.2026).