


Scientific journal
Fundamental research
ISSN 1812-7339
"Перечень" ВАК
ИФ РИНЦ = 1,798
ROLE OF TASK-1/TASK-3 TYPES OF BACKGROUND 2P POTASSIUM CHANNELS IN THE BRIEF HYPOXIA INDUCED CHANGES IN THE ACTIVITY OF HIPPOCAMPAL CA1 PYRAMIDAL NEURONS

Известно, что длительная непрерывная гипоксия является одной из главных причин необратимого повреждения клеток мозга [12]. В отличие от длительной гипоксии повторяющиеся кратковременные гипоксические эпизоды, не вызывая необратимого повреждения пирамидных нейронов поля СА1 гиппокампа мозга крысы, способны индуцировать в этих нейронах такие долговременные формы функциональной нейропластичности, как быстрое гипоксическое прекондиционирование и устойчивая постгипоксическая гипервозбудимость нейронов [3, 7].
Наши предварительные эксперименты показали, что Са2+- и АТФ-зависимые мембранные калиевые каналы, являющиеся одними из основных мишеней действия гипоксии, играют важную роль в механизмах индуцируемых кратковременными эпизодами гипоксии перестройках в активности пирамидных нейронов поля СА1 гиппокампа. В частности, Са2+-активирумые калиевые каналы большой проводимости играют важную роль как в механизмах быстрого гипоксического прекондиционирования, так и развитии постгипоксической гипервозбудимости [7]. С другой стороны, АТФ-зависимые калиевые каналы, относящиеся к подтипу митохондриальных КАТФ каналов, достоверно не влияют на развитие постгипоксической гипервозбудимости, но также играют важную роль в механизмах быстрого гипоксического прекондиционирования в пирамидных нейронах поля СА1 гиппокампа [9].
Относительно недавно было обнаружено, что плазматическая мембрана различных нейронов мозга, в том числе и пирамидных нейронов поля СА1 гиппокампа, содержит особый тип калиевых каналов, субъединицы которых формируют четыре трансмембранные группировки и две трансмембранные поры (фоновые 2Р калиевые каналы) [6]. Эти каналы селективны для ионов калия, а их функция связана с поддержанием мембранного потенциала клетки в покое, а также регуляцией гомеостаза К+ и фоновой возбудимости нейронов. Результаты исследований показывают, что через фоновые 2Р калиевые каналы осуществляют свои модулирующие эффекты некоторые гормоны и нейромедиаторы, анестетики, полиненасыщенные жирные кислоты, а также такие физико-химические факторы, как гипоксия, изменения вне- и внутриклеточного рН, температура и механическое растяжение клеточной мембраны [6]. Эти калиевые каналы являются потенциал-независимыми, неинактивирующимися и постоянно-активными. Их блокада вызывает деполяризацию, а активация – гиперполяризацию мембраны нейронов. На основе анализа структуры и функциональных свойств семейство 2Р калиевых каналов млекопитающих подразделяется на 6 подсемейств, включающих 15 членов. В частности, в гиппокампе экспрессируются такие члены семейства 2Р калиевых каналов, как TASK, TREK, TRAAK и TWIK. Отмечается, что в поле СА1 гиппокампа крысы TASK тип каналов экспрессируется только в пирамидных клетках, но не интернейронах [14]. Однако в экспериментах на мышах было обнаружено, что данный тип каналов выявляется также в 30–50 % интернейронов stratum oriens поля СА1 гиппокампа [15].
В связи с тем, что TASK-1/TASK-3 типы фоновых 2Р калиевых каналов чувствительны к гипоксии и существенным образом влияют на возбудимость нейронов, целью настоящей работы было исследование влияния селективного блокатора этих каналов А-1899 [13] на индуцируемые кратковременными эпизодами гипоксии изменения в активности нейронов гиппокампа непосредственно во время эпизода гипоксии, прекондиционирующее действие первых двух эпизодов гипоксии на третий и постгипоксическую гипервозбудимость исследуемых нейронов.
Материалы и методы исследования
Работа осуществлялась с соблюдением принципов гуманного обращения с животными (директива Европейского сообщества 86/609/EC). В экспериментах использовали двухмесячных самцов крыс Вистар (n = 18). Срезы гиппокампа толщиной 250–300 мкм помещались в камеру погруженного типа и суперфузировались раствором Кребса–Рингера следующего состава (мМ): NaCl – 124, KCl – 3, MgSO4 – 2, CaCl2 – 2, NaHCO3 – 26, KH2PO4 – 1.24, глюкоза – 10, O2/CO2 – 95/5 % (pH = 7,4), при температуре раствора 32 °С и скорости его протока 2,4 мл/мин. Регистрацию активности начинали через 2,5 часа от начала суперфузии. Популяционные спайки (ПС) регистрировались от пирамидных нейронов поля СА1 гиппокампа в ответ на электрическую тестирующую стимуляцию коллатералей Шаффера до, во время и после предъявления трех 3-минутных эпизодов гипоксии, разделенных 10-минутными интервалами. На рисунке, а показаны расположение гиппокампа в мозге крысы, схема среза гиппокампа с локализацией регистрирующего и стимулирующего электродов, форма регистрируемых ПС ответов при нормальных условиях (до предъявления эпизодов гипоксии) и трансформация формы ПС ответов через 60 мин после предъявления эпизодов гипоксии. Эффективность гипоксического эпизода снижать амплитуду ПС оценивалась по следующей формуле:
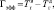
где Гэфф – эффективность эпизода гипоксии снижать амплитуду ПС (в секундах); 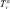 – время, через которое амплитуда ПС депрессируется до 50 % от ее прегипоксических значений от начала эпизода гипоксии;
– время, через которое амплитуда ПС депрессируется до 50 % от ее прегипоксических значений от начала эпизода гипоксии;  – время, через которое амплитуда ПС восстанавливается до 50 % от ее прегипоксических значений после окончания действия гипоксии. Оценка развития постгипоксической гипервозбудимости нейронов в срезах гиппокампа, проявляющейся в форме эпилептиформных ПС-разрядов, производилась по разработанной в нашей лаборатории оригинальной методике [2, 3]. Измерялось число ПС в разряде (ЧПС) на стимул, интенсивность которого приводила к появлению второго ПС в ответе. Быстрое гипоксическое прекондиционирование оценивалось по формуле
– время, через которое амплитуда ПС восстанавливается до 50 % от ее прегипоксических значений после окончания действия гипоксии. Оценка развития постгипоксической гипервозбудимости нейронов в срезах гиппокампа, проявляющейся в форме эпилептиформных ПС-разрядов, производилась по разработанной в нашей лаборатории оригинальной методике [2, 3]. Измерялось число ПС в разряде (ЧПС) на стимул, интенсивность которого приводила к появлению второго ПС в ответе. Быстрое гипоксическое прекондиционирование оценивалось по формуле
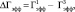
где 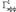 и
и  представляют собой значения Гэфф соответственно для первого и третьего эпизодов гипоксии. Электрическая активность нейронов измерялась на серии из 10 одиночных стимулов градуально увеличивающейся интенсивности. Частота импульсов в серии была 0,1 Гц, частота серий – 1 серия/10 мин. Гипоксические эпизоды предъявлялись без добавления в среду А-1899 или через 10 мин после его добавления в концентрации 0,5 мкМ, при которой происходила блокада как TASK-1, так и TASK-3 каналов [13]. А-1899 (фирма Santa Cruz, USA) растворялся в растворе DMSO, концентрация которого в суперфузирующем растворе (0,05 %) не влияла на параметры активности нейронов. Время приложения к срезам гиппокампа А-1899 составляло 40 мин. В отдельной серии экспериментов А-1899 добавлялся к срезам гиппокампа в отсутствии эпизодов гипоксии.
представляют собой значения Гэфф соответственно для первого и третьего эпизодов гипоксии. Электрическая активность нейронов измерялась на серии из 10 одиночных стимулов градуально увеличивающейся интенсивности. Частота импульсов в серии была 0,1 Гц, частота серий – 1 серия/10 мин. Гипоксические эпизоды предъявлялись без добавления в среду А-1899 или через 10 мин после его добавления в концентрации 0,5 мкМ, при которой происходила блокада как TASK-1, так и TASK-3 каналов [13]. А-1899 (фирма Santa Cruz, USA) растворялся в растворе DMSO, концентрация которого в суперфузирующем растворе (0,05 %) не влияла на параметры активности нейронов. Время приложения к срезам гиппокампа А-1899 составляло 40 мин. В отдельной серии экспериментов А-1899 добавлялся к срезам гиппокампа в отсутствии эпизодов гипоксии.
Ввод вызванных электрических ответов в компьютер, а также их обработка в режиме off-line осуществлялись с помощью соответствующего программного обеспечения. Данные представлялись в виде средняя ± ошибка средней. Оценку достоверности влияния исследуемых веществ на индуцируемые гипоксией изменения в регистрируемых параметрах осуществляли по критерию Стьюдента: непарный t-тест для одних и тех же моментов времени регистрации активности в «гипоксических» и «гипоксических + А1899» срезах гиппокампа и парный t-тест при сравнении пре- и постгипоксических значений амплитуды ПС. При значениях P ≤ 0,05 различия считались достоверными.
Результаты исследований и их обсуждение
В концентрации 0,5 мкМ А-1899 не влиял на амплитуду ПС, регистрируемых от исследуемых нейронов. Однако этот селективный блокатор TASK-1/TASK-3 каналов достоверно снижал эффективность угнетающего действия гипоксии на амплитуду ПС во время эпизода (рисунок, б). Значение Гэфф для первого эпизода гипоксии на фоне А-1899, равное 292 ± 19 с (n = 5), достоверно (Р = 0.03) отличалось от аналогичного значения Гэфф для первого эпизода гипоксии без добавления А-1899 (350 ± 12 с, n = 8). Известно, что кратковременный эпизод гипоксии вызывает в пирамидных нейронах поля СА1 гиппокампа начальную гиперполяризацию в течение первой минуты гипоксии, устраняющую спонтанную и вызванную активность нейронов, и последующую градуально увеличивающуюся деполяризацию мембранного потенциала [4, 5, 10]. Начальная гиперполяризация, связанная с увеличением мембранной проводимости к ионам калия, приводит к накоплению этих ионов во внеклеточной среде и нарушению их трансмембранного градиента. Известно, что в поддержании мембранного потенциал покоя нейронов основная роль принадлежит фоновым 2Р калиевым каналам, причем в отличие от других типов этих каналов TASK-1/TASK-3 каналы чувствительны к гипоксии [6]. По-видимому, устранение вызванного эпизодом гипоксии нарушения трансмембранного градиента ионов калия блокатором TASK-1/TASK-3 калиевых каналов способствует меньшему нарушению градиента этих ионов и более быстрому восстановлению амплитуды ПС после окончания действия кратковременной гипоксии.
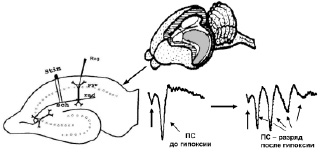
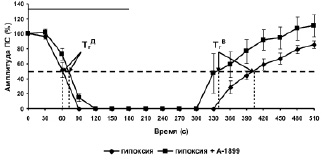
Влияние А-1899 на угнетающее действие трëхминутного эпизода гипоксии на амплитуду ПС: а – схема расположения стимулирующего (Stim) и регистрирующего (Reg) электродов в срезе гиппокампа и типичные формы ПС ответов до и после трёх 3-минутных эпизодов гипоксии. Sch – коллатерали Шаффера; rad – радиальный слой; pyr – пирамидный слой; б – динамика изменения амплитуды ПС во время эпизода гипоксии и гипоксии на фоне А-1899 (0,5 мкМ). По оси абсцисс – время в секундах, по оси ординат – амплитуда ПС в процентах от ее прегипоксических значений. Сплошная линия на графике – время приложения эпизода гипоксии или эпизода гипоксии с А-1899. Пунктирная линия отмечает 50 % снижение и 50 % восстановление амплитуды ПС после начала и окончания эпизода гипоксии
На фоне А-1899 (0,5 мкМ) наблюдалась также тенденция к снижению прекондиционирующего действия первых двух эпизодов гипоксии на эффективность третьего эпизода подавлять амплитуду ПС. Однако это снижение было недостоверным. Известно, что прекондиционирующие эффекты развиваются в двух временных диапазонах: в течение нескольких минут (быстрое прекондиционирование) и нескольких суток (задержанное прекондиционирование) [11]. Как показали наши предыдущие эксперименты, в механизмах быстрого гипоксического прекондиционирования важную роль играют митохондриальные АТФ-зависимые К+ каналы (9) и Са2+-активируемые К+ каналы большой проводимости [7]. Результаты настоящего исследования продемонстрировали, что TASK-1/TASK-3 типы фоновых 2Р калиевых каналов, по-видимому, не играют существенной роли в этих механизмах.
Анализ влияния А-1899 на развитие постгипоксической гипервозбудимости показал, что в концентрации 0,5 мкМ он не влияет на ее развитие. При нормальных физиологических условиях пирамидные нейроны поля СА1 гиппокампа генерируют одиночные потенциалы действия. Однако при определенных патологических условиях, в частности, после гипоксии, ишемии или действия потенциально эпилептогенных факторов эти нейроны способны генерировать пачки потенциалов действия. Генерация такой индуцируемой гипоксией пачечной активности (постгипоксическая гипервозбудимость) может быть связана как с входящими кальциевыми токами через потенциал- и лиганд-регулируемые мембранные каналы [3], так и входящим натриевым током через медленно-инактивирующиеся натриевые каналы [1]. Кроме того, наши предыдущие исследования показали, что в отличие от АТФ-зависимых калиевых каналов кальций-активируемые калиевые каналы также играют важную роль в развитии постгипоксической гипервозбудимости [7, 8]. Результаты настоящей работы позволяют сделать заключение, что чувствительные к гипоксии TASK-1/TASK-3 фоновые 2Р калиевые каналы не играют существенной роли в развитии постгипоксической гипервозбудимости.
Заключение
Результаты проведенных исследований позволяют заключить, что фоновые 2Р калиевые каналы TASK-1/TASK-3 типов играют важную роль в механизмах угнетающего действия гипоксии на активность пирамидных нейронов поля СА1 гиппокампа во время гипоксии, но не вносят существенного вклада в такие индуцируемые кратковременными эпизодами гипоксии формы функциональной нейропластичности, как быстрое гипоксическое прекондиционирование и развитие устойчивой постгипоксической гипервозбудимости.
Работа выполнена при поддержке Российского фонда фундаментальных исследований (проект № 11-04-00013а).
Рецензенты:
Зинченко В.П., д.б.н., профессор, заведующий лабораторией внутриклеточной сигнализации, ФГБУН «Институт биофизики клетки Российской академии наук», г. Пущино;
Журавлева З.Н., д.б.н., в.н.с., лаборатория системной организации нейронов, ФГБУН «Институт теоретической и экспериментальной биофизики Российской академии наук», г. Пущино.
Работа поступила в редакцию 06.11.2013.
Библиографическая ссылка
Левин С.Г., Конаков М.В., Годухин О.В. РОЛЬ ФОНОВЫХ 2Р КАЛИЕВЫХ КАНАЛОВ TASK-1/TASK-3 ТИПОВ В ИНДУЦИРУЕМЫХ КРАТКОВРЕМЕННЫМИ ЭПИЗОДАМИ ГИПОКСИИ ИЗМЕНЕНИЯХ В АКТИВНОСТИ ПИРАМИДНЫХ НЕЙРОНОВ ПОЛЯ СА1 ГИППОКАМПА // Фундаментальные исследования. 2013. № 11-1. С. 104-107;URL: https://fundamental-research.ru/en/article/view?id=33018 (дата обращения: 05.07.2026).