


Scientific journal
Fundamental research
ISSN 1812-7339
"Перечень" ВАК
ИФ РИНЦ = 1,798
CHARACTERIZATION OF RHIZOSPHERA´S AND RHIZOPLANA´S MICROBIAL COMMUNITY OF CUCURBITA PEPO L.

Положительная роль микроорганизмов в выращивании сельскохозяйственных культур связана с образованием симбиотических отношений между растением и микробным сообществом в ризосфере растений [1, 7]. В образуемом эктосимбиозе корневые экссудаты растений являются субстратом и факторами роста некоторых групп микробных сообществ, которые выполняют роль антифитопатогенов, утилизаторов нежелательных продуктов метаболизма растений, регуляторов общей концентрации микроорганизмов в почве, регуляторов подвижности и кругооборота минеральных веществ в агроэкосистеме [5].
К настоящему времени накоплен большой экспериментальный материал, доказывающий значительную роль ризобактерий в жизнедеятельности многих сельскохозяйственных растений [2, 8, 9]. Однако сведения о ризосферной микрофлоре Cucurbita pepo L., являющейся ценной овощной, кормовой и масличной культурой, отсутствуют.
Цель: изучить видовой состав микробного сообщества ризосферы и ризопланы тыквы в различные периоды роста и развития.
Материал и методы исследования
Объектом исследования явились микроорганизмы, выделенные из ризосферы и ризопланы тыквы обыкновенной (Cucurbita pepo L.) Оценку видового состава изучаемого микробоценоза проводили методом посева полученной суспензии на плотные питательные среды: МПА, Эндо, Симмонса, культуры инкубировали при t = 37 °С в течение 48 ч [6]. Исследования проводили в течение четырёх вегетационных периодов: фазе всходов, бутонизации, цветения и плодоношения. Идентификацию микроорганизмов осуществляли на основе изучения морфологических, тинкториальных, культуральных, физиолого-биохимических свойств выделенных микроорганизмов с помощью программного обеспечения для автоматизированной идентификации бактерий производства ООО «НПО Диагностические системы».
Результаты исследования и их обсуждение
Исследования показали, что микробное сообщество прикорневой зоны Cucurbita pepo характеризуется высокой плотностью и видовым разнообразием. При этом качественный и количественный состав микробоценоза ризосферы зависит от фазы развития растения и меняется в течение всего вегетационного периода.
В фазе всходов были выделены бактерии р. Pseudomonas и Enterobacter spp. Бациллы на первых фазах развития растения распространены слабо, так как эти бактерии плохо утилизируют простые органические соединения, синтезирующиеся в начальные периоды вегетации. Ризосферный эффект увеличивался после прорастания семени и достигал максимума в период цветения и плодоношения растений. В стадии бутонизации, цветения и плодоношения были выделены: Pseudomonas fluorescens, Enterobacter spp., неферментирующие бактерии (Acinetobacter haemoliticus и Shewanella putrefaeciens), энтеробактерии (Citrobacter freundi, Serratiae odorivera, E. amnigenes, E. cancerogenes, E. intermedius, E. gergoviae. E. aerogenes) и спорообразующие бактерии р. Bacillus (B. subtilis, B. cereus, B. megaterium) (таблица).
Видовой состав микробоценоза ризосферы Cucurbita pepo в различные периоды вегетации
|
Виды ризобактерий |
Фазы вегетации |
|||
|
всходов |
бутонизации |
цветения |
плодоношения |
|
|
P. fluorescens |
+ |
+ |
– |
– |
|
Enterobacter spp. |
+ |
+ |
– |
– |
|
E. amnigenes |
– |
– |
+ |
– |
|
E. cancerogenes |
– |
– |
+ |
– |
|
E. aerogenes |
– |
– |
– |
+ |
|
E. intermedius |
– |
– |
– |
+ |
|
E. gergoviae |
– |
– |
– |
+ |
|
Citrobacter freundi |
– |
– |
– |
+ |
|
Serratiae odorivera |
– |
– |
– |
+ |
|
Acinetobacter haemoliticus |
– |
– |
+ |
– |
|
Shewanella putrefaeciens |
– |
– |
– |
+ |
|
Bacillus subtilis |
– |
+ |
– |
+ |
|
B. cereus |
– |
+ |
– |
– |
|
B. megaterium |
– |
– |
– |
+ |
Из таблицы видно, что наибольшее видовое разнообразие микробного сообщества ризосферы было отмечено в фазе цветения и плодоношения. Такая смена качественного состава микробного сообщества ризосферы, по-видимому, связана с изменением синтетической активности корней растений на различных стадиях вегетации: в начальные периоды преобладают бактерии, питающиеся корневыми выделениями растений, в более поздние периоды доминируют гидролитики, разлагающие растительный опад.
В ходе изучения квантитативных показателей микробного сообщества ризосферы было установлено, что общая численность ризобактерий изменяется в зависимости от фазы развития Cucurbita pepo. В фазе всходов общая численность ризобактерий составляла 2,56 ± 0,035 lg КОЕ/мл, из них 95 % приходилось на неспорообразующие бактерии. В фазе бутонизации плотность колонизации микробного сообщества ризосферы составляла 2,97 ± 0,023 lg КОЕ/мл, из них на долю неспорообразующих форм бактерий приходилось 86 %. В фазе цветения и плодоношения общая численность ризобактерий составляла 3,54 ± 0,031 lg и 3,61 ± 0,042 lg КОЕ/мл соответственно. На поздних фазах вегетации (цветения и плодоношения) доля спорообразующих бацилл от общей численности ризобактерий составляла 13 и 48 % соответственно.
Известно, что с наступлением фазы плодоношения происходит снижение метаболической деятельности корней, что проявляется в снижении количества выделяемых метаболитов и связано с уменьшением снабжения корневой системы углеводами [3, 4]. Таким образом, доля спорообразующих бацилл в общей численности ризобактерий увеличивается в процессе роста и развития Cucurbita pepo и максимальна в фазе плодоношения.
Микрофлора ризоплана в определенной степени также отличается от микробного ценоза ризосферы. В ризоплане обычно преобладают грамотрицательные бактерии, в то время, как ризосферу населяют в основном спорообразующие бактерии. Доминирующее место в микробном ценозе ризосферы тыквенных растений занимали Citrobacter freundii, их численность достигала 3,2 ± 0,001 lg КОЕ/мл, количественные показатели B. subtilis и B. megaterium составили 3,1 ± 0,012 и 2,9 ± 0,014 lg КОЕ/мл соответственно. Непосредственно на корнях растений выявлено достоверно меньшее количество микроорганизмов (р < 0,05), чем в прикорневой зоне.
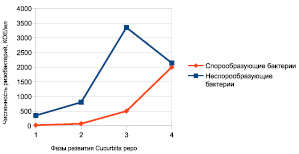
Рис. 1. Динамика численности ризобактерий Cucurbita pepo в различные периоды вегетации (1– фаза всходов, 2 – фаза бутонизации, 3 – фаза цветения, 4 – фаза плодоношения)
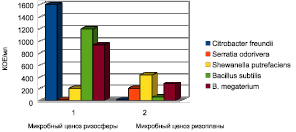
Рис. 2. Квантитативные показатели микробного ценоза ризосферы и ризопланы Cucurbita pepo
Микробное сообщество ризоплана было представлено в основном грамотрицательными палочками Shewanella putrefaeciens и Serratia freundii, их количество достигало 2,6 ± 0,015 и 2,3 ± 0,011 lg КОЕ/мл соответственно. Квантитативные показатели B.megaterium и B.subtilis в микробоценозе ризопланы составили 2,4 ± 0,015 и 1,8 ± 0,011 lg КОЕ/мл соответственно (р < 0,05).
Выводы
1. Видовой состав микробного сообщества ризосферы и ризопланы имеет существенные отличия. Микрофлора ризосферы представлена спорообразующими бактериями B. subtilis и B. megaterium, энтеробактериями Citrobacter freundi и Shewanella putrefaeciens. В микробном населении ризопланы доминируют грамотрицательные бактерии Shewanella putrefaeciens и Serratia odorivera, в достоверно меньшем количестве выделены B. subtilis и B. Megaterium.
2. Видовой состав микробного сообщества ризосферы и ризопланы тыквы изменяется в зависимости от периода роста и развития. В фазе всходов микробоценоз представлен Pseudomonas spp. и Enterobacter spp., в стадии бутонизации – Pseudomonas fluorescens, Enterobacter spp., Bacillus subtilis, B. cereus. В стадии цветения и плодоношения были выделены неферментирующие бактерии: Acinetobacter haemoliticus и Shewanella putrefaeciens, представители семейства энтеробактерий C. freundi, S. odorivera, E. amnigenes, E. cancerogenes, E. intermedius, E. gergoviae. E. aerogenes, и спорообразующие бактерии B. subtilis, B. Megaterium.
3. Квантитативные показатели микробного сообщества ризосферы и ризоплана имеют существенные отличия. Наиболее плотнозаселенной нишей является ризосфера, где доминирующее место занимает Citrobacter freundii и спорообразующие бактерии – B. subtilis и B. megaterium. Плотность колонизации бактерий в ризоплане достоверно меньше, основная доля приходилась на грамотрицательные бактерии Shewanella putrefaeciens и Serratia odorivera. Количественные показатели B. subtilis, B. megaterium были достоверно меньше, чем в ризосфере (1,8 ± 0,011 и 2,4 ± 0,015 lg КОЕ/мл соответственно; р < 0,05).
4. Общая численность ризобактерий Cucurbita pepo зависит от фазы роста и развития растения. В фазе всходов общая численность ризобактерий составляла 2,56 ± 0,035 lg КОЕ/мл, из них 95 % приходилось на неспорообразующие бактерии и 5 % – на спорообразующие бациллы. В фазе бутонизации общая численность бактерий ризосферы составляла 2,97 ± 0,023 lg КОЕ/мл, из них на долю неспорообразующих микробов и бацилл приходилось 86 и 14 % соответственно. В фазе цветения на долю спорообразующих бактерий приходилось 13 % от общей численности бактерий, равной 3,54 ± 0,031 lg КОЕ/мл. Максимум численности бацилл был отмечен в фазе плодоношения и составлял 48 % от общей численности ризобактерий, равной 3,61 ± 0,042 lg КОЕ/мл.
Рецензенты:
Золотухин В.В., д.б.н., профессор кафедры зоологии, ФГБОУ ВПО «Ульяновский государственный педагогический университет им. И.Н. Ульянова», г. Ульяновск;
Артемьева Е.А., д.б.н., профессор кафедры зоологии, ФГБОУ ВПО «Ульяновский государственный педагогический университет им. И.Н. Ульянова», г. Ульяновск.
Работа поступила в редакцию 05.12.2013.
Библиографическая ссылка
Артамонова М.Н., Потатуркина- Нестерова Н.И. ХАРАКТЕРИСТИКА МИКРОБНОГО СООБЩЕСТВА РИЗОСФЕРЫ И РИЗОПЛАНА CUCURBITA PEPO L. // Фундаментальные исследования. 2013. № 10-14. С. 3067-3070;URL: https://fundamental-research.ru/en/article/view?id=32960 (дата обращения: 26.07.2026).