


Scientific journal
Fundamental research
ISSN 1812-7339
"Перечень" ВАК
ИФ РИНЦ = 1,798
CONTRIBUTION OF MYCOGENIC WOOD DESTRUCTION ON FORMATION OF MICROCLIMATE OF THE SOUTHERN PREURALS FORESTS

Биогеоценозы, в частности, лесные биогеоценозы, являются сложными природными образованиями, включающими в себя как биотические, так и абиотические компоненты. В биогеоценозе формируется специфическая биотическая среда, параметры которой изначально зависят от многих физико-географических условий, а в дальнейшем, по мере формирования и развития биогеоценоза, испытывают все более существенное влияние со стороны биотических компонентов. Организмы, населяющие биогеоценоз, с одной стороны, вынуждены «принимать» существующие в нем условия, а с другой – они активно участвуют в средообразовании в самых разных его аспектах (формирование почвы, микрорельефа, микроклимата и т.д.).
Влияние организмов на формирование среды своего обитания, особенно в плане формирования микроместообитаний с микроклиматическим статусом, наиболее соответствующим этим организмам, а также оценка вклада этих обитаний в формирование общей биогеоценотической среды относительно слабо изучено. На существование в биогеоценозах разных микроклиматических условий и микроместообитаний и их внешнее проявление – неравномерность пространственного распределения организмов в лесу и их функционрирования указывали многие авторы [4, 5, 7, 9 и др.]. Обычно это качество биогеоценоза рассматривается как мозаичность, т.е. своеобразное сочетание в его пределах отдельных структурных единиц (консорций [3, 11 и др.] или тессер – элементарных единиц лесных биогеоценозов, на уровне которых реализуются взаимосвязи растительность – почвенная биота – почва [8, 10]).
Если говорить о биогеоценозе с консортивной точки зрения, каждая из консорций, занимая определенное место в экотопе и обладая своеобразным комплексом организмов, вносит собственную лепту в формирование мозаичности микроклиматических условий биогеоценоза. Не являются исключением и консорции, ядром которых выступают не живые деревья, а древесные остатки (валежные стволы, пни и т.п.) (существование таких консорций признается рядом авторов [1, 2, 6, 11 и др.]. Эти консорции априори должны обладать специфическим микроклиматом, поскольку древесина, подверженная микогенной деструкции, активно выделяет углекислый газ, и гниение должно сопровождаться особым режимом температуры и влажности.
Материалы и методы исследования
Для выяснения особенностей формирования микроклимата в консорциях, формируемых деструктурируемой древесиной, нами были проведен суточный мониторинг показателей влажности и температуры в пределах данных консорций в лесах предгорий Южного Урала (Троицкий заказник Тюльганского района Оренбургской области). Исследования проводились в июне-июле 2013 года. Мониторинг температуры и влажности производился с интервалом в 10 минут с использованием логгеров EClerk-USB-RHT. Логгеры закладывались в трехкратной повторности в березняке разнотравном, осиннике разнотравном и в сосновой посадке в разных локациях – на высоте 1,5 м, около разлагающихся древесных стволов, непосредственно внутри разлагающейся древесины, а также в плодовых телах трутовика настоящего (Fomes fomentarius (L.: Fr.) Fr.). Результаты обрабатывались при помощи пакета статистического анализа программы MS Excel (корреляционный анализ).
Результаты исследования и их обсуждение
Анализ суточной динамики температур в березняке разнотравном показал наличие двух достаточно явно различающихся тенденций динамики температуры: одна из них характерна для среды биогеоценоза в целом, а вторая характерна для деструктурируемого субстрата и плодовых тел трутовиков (рис. 1).
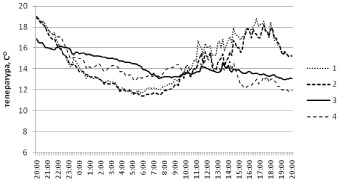
Рис. 1. Суточная динамика температуры в березняке разнотравном. Локации логгеров: 1 – среда; 2 – вблизи валежного ствола березы; 3 – внутри валежного ствола березы; 4 – внутри плодового тела трутовика настоящего на валежном стволе березы
Динамика температуры в первом случае явно зависит от освещенности биогеоценоза, о чем свидетельствуют максимальные показатели, приходящиеся на период времени с 12 до 19 часов дня. Корреляция между динамикой температур в разных частях биогеоценоза при этом достигает 0,98.
Вторая из отмеченных тенденций характеризуется отсутствием выраженного, учитываемого влияния освещенности биотопа. Корреляция между ходом температур внутри валежных стволов березы и внутри плодовых тел трутовиков достигает 0,85. При этом более постоянный уровень температуры характерен для разлагающейся древесины. Температура плодовых тел грибов наиболее существенно снижается в самое жаркое время суток, что, вероятно, обеспечивается значительным повышением испарения.
В отношении динамики влажности аналогичные тенденции статистически менее выражены, хотя графический анализ (рис. 2) показывает, что для разлагаемого субстрата и плодового тела трутовика свойственна более высокая влажность и меньшее ее варьирование в течение суток.
Помимо отдельных показателей влажности и температуры в анализе был учтен гидротермический коэффициент (отношение влажности к температуре) как интегральный показатель температурно-влажностных условий, отражающий один из наиболее специфичных показателей среды – испаряемость.
Указанные выше различия в динамике показателей наиболее ярко проявились именно при анализе гидротермического коэффициента (рис. 3).
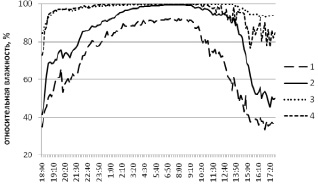
Рис. 2. Суточная динамика влажности в березняке разнотравном. Локации логгеров: 1 – среда; 2 – вблизи валежного ствола березы; 3 – внутри валежного ствола березы; 4 – внутри плодового тела трутовика настоящего на валежном стволе березы
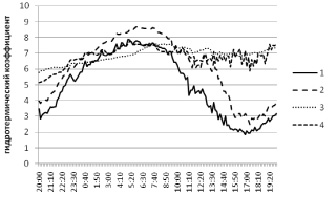
Рис. 3. Суточная динамика гидротермического градиента в березняке разнотравном. Локации: 1 – среда; 2 – вблизи валежного ствола березы; 3 – внутри валежного ствола березы; 4 – внутри плодового тела трутовика настоящего на валежном стволе березы
Сходные тенденции отличия в режиме микроместообитаний у древесных остатков, подверженных микогенной деструкции, были отмечены нами и в отношении лесных сообществ с другими лесообразующими видами.
В осиннике разнотравном нами производился мониторинг климатических показателей как на субстрате с плодовыми телами, так и на субстрате со скрытой гнилью. В обоих случаях тенденции динамики показателей в целом аналогичны отмеченным в березняке (рис. 4, 5).
Подводя итого анализу приведенных данных, можно сделать вывод, что микогенная деструкция древесины, изменяя климатические показатели микроместообитаний, вносит определенный вклад в формирование климатических показателей биогеоценоза. Влияние деструктурируемых единиц субстрата на микроклиматические показатели заключается в смягчении перепадов температуры и влажности, обусловленных их колебаниями во внешней среде. Это сглаживание резких изменений можно, вероятно, объяснить рядом причин – изменением структуры древесины, снижающим ее влагопотерю; динамичностью испаряемости влаги из плодового тела гриба как защитной реакцией на перегрев.
Экологическое значение создания особых микроклиматических условий вблизи деструктурируемого грибами субстрата, вероятно, заключается в создании по возможности оптимальных условий для дальнейшей микогенной деструкции древесины и, соответственно, дальнейшего роста и развития гриба. Кроме того, можно предположить, что данный микроклимат благоприятен для заселения близлежащих субстратов грибами. Тем самым микогенная деструкция на одном субстрате провоцирует более активное заселение грибами соседних субстратов. Однако для точного установления этого факта необходимо проведение дополнительных исследований.
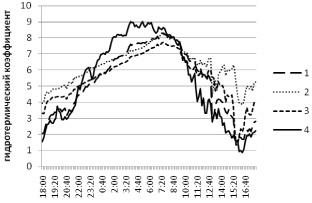
Рис. 4. Суточная динамика гидротермического градиента в осиннике разнотравном на стволе с плодовыми телами Fomes fomentarius (L.: Fr.) Fr. Локации логгеров: 1 – возле валежного ствола осины; 2 – внутри валежного ствола осины; 3 – внутри плодового тела трутовика настоящего на валеже осины; 4 – среда
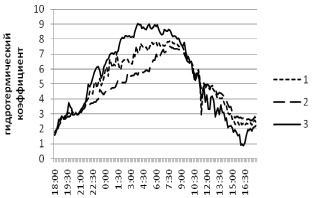
Рис. 5. Суточная динамика гидротермического градиента в осиннике разнотравном на стволе со скрытой гнилью. Локации логгеров: 1 – внутри сухостойного ствола осины; 2 – на сухостойном стволе осины; 3 – среда
Рецензенты:
Русанов А.М., д.б.н., профессор, заведующий кафедрой общей биологии, ФГБОУ ВПО «Оренбургский государственный университет», г. Оренбург;
Паршина Т.Ю., д.б.н., доцент, профессор кафедры зоологии, экологии и анатомии, ФГБОУ ВПО «Оренбургский государственный педагогический университет», г. Оренбург.
Работа поступила в редакцию 03.10.2013.Библиографическая ссылка
Сафонов М.А., Булгаков Е.А. ВКЛАД МИКОГЕННОЙ ДЕСТРУКЦИИ ДРЕВЕСИНЫ В ФОРМИРОВАНИЕ МИКРОКЛИМАТА ЛЕСОВ ЮЖНОГО ПРИУРАЛЬЯ // Фундаментальные исследования. 2013. № 10-12. С. 2674-2678;URL: https://fundamental-research.ru/en/article/view?id=32850 (дата обращения: 20.06.2026).