


Scientific journal
Fundamental research
ISSN 1812-7339
"Перечень" ВАК
ИФ РИНЦ = 1,798
THE ROLE OF PHOSPHOLIPID ENVIRONMENT IN THE MEMBRANE PROTEIN TRANSLOCATION IN E.COLI

Примерно 20 % около 4000 белков Escherichia coli являются мембранными белками [1]. Процесс встраивания мембранных белков в мембрану опосредован работой специальных белковых комплексов, формирующих канал. Считается, что только некоторые белки с малой молекулярной массой обладают способностью к спонтанному встраиванию, тогда как подавляющее большинство задействует транслоказы и инсертазы. Различают ко-трансляционный механизм встраивания, при котором фолдинг белка осуществляется в процессе встраивания, и пост-трансляционный, в процессе которого происходит транслокация уже свернутого в нативную структуру белка [2]. Известны как минимум две системы транслокации у E.coli: Tat-система и комплекс SecYEG.
Tat-система является менее изученной по сравнению с комплексом SecYEG, однако известно, что транслокация белками Tat происходит по пост-трансляционному механизму, то есть готовые к выполнению функций белки с нативной структурой транспортируются через мембрану. Tat-транслоказа (twin arginine translocation) содержит белки TatA, TatB и TatC. Белки TatA (молекулярная масса 9 кДа), TatB (18 кДа) имеют по одному трансмембранному домену, N-концевой сегмент располагается внутри цитоплазмы, C-конец ориентирован в сторону периплазматического пространства. Белок TatC содержит 6 трансмембранных доменов, N- и C-концевые сегменты расположены внутри цитоплазмы. Обнаруживают 2 крупных белковых комплекса, содержащих Tat-белки при солюбилизации мембраны с помощью детергентов. Первый комплекс состоит из TatA и относительно небольшого количества TatB, в то время как другой содержит почти эквимолярное количество TatB и TatC белков. Оба комплекса достигают молекулярной массы свыше 600 кДа [3].
TatC является ответственным за распознавание сигнальной последовательности S/TRRxFLK, всегда содержащей 2 аргинина, участвующих в механизме транслокации и давших название данной системе. Длина сигнальной последовательности Tat-системы в среднем равна 38 аминокислотам, в то время как для Sec-транслокации длина составляет 24 аминокислоты. В последовательности, распознаваемой Tat-системой, присутствует полярная область, по всей видимости, препятствующая взаимодействию с белками Sec-системы. Сигнальная последовательность удаляется сигнальной пептидазой непосредственно после транслокации. Считается, что TatA начинает формировать трансмембранный канал по мере связывания TatC с сигнальной последовательностью, при этом процесс формирования канала является зависимым от градиента мембранного потенциала. Образуется комплекс TatBC-TatA, который носит динамичный характер и распадается сразу после транспорта субстрата. Существуют сведения о том, что белок TatA обладает двойной топологией в мембране [4].
Существует ряд белков, выполняющих функцию контроля качества фолдинга транслоцируемых белков. Цитоплазматические шаперон-подобные белки с рядом ко-факторов взаимодействуют с синтезируемой полипептидной цепью для предотвращения преждевременного связывания сигнальной последовательности с Tat-транслоказой. Данный тип шаперонов получил название REMP (redox enzyme maturation protein) у E.coli, гомологи которого обнаружены и в других организмах [5].
Структура и функции транслокона SecYEG E. coli
Основной системой секреции белков у E.coli является Sec-транслокон, выполняющий транспорт через мембрану или встраивание интегральных белков. Процесс транспорта мембранного белка к месту расположения в мембране начинается с мечения белка и его распознавания. Мечение мембранного белка происходит на достаточно раннем этапе его синтеза – до момента покидания полипептидной цепи рибосомы. Синтезируемая полипептидная цепь взаимодействует своим гидрофобным сегментом с сигнал-распознающей частицей SRP (от signal recognition particle) [6]. Бактериальная SRP состоит из белкового компонента Ffh и 4,5S РНК. Взаимодействующий сегмент, как правило, является первым трансмембранным доменом, однако может также быть удаленным от первого домена и даже не являться встраиваемым сегментом (с таким белком работает в дальнейшем инсертаза KdpD) [7].
Далее SRP с полипептидной цепью формирует комплекс с сигнальным рецептором SR, функцию которого в клетках E.coli выполняет белок FtsY, расположенный на мембране. Дезагрегация комплекса и высвобождение целевого белка требует гидролиза ГТФ. Как SRP, так и FtsY изначально имеют связь с ГТФ, гидролиз которого помогает сформировать комплекс благодаря непосредственному взаимодействию с их NG-доменами. Ffh и FtsY имеют 2 гидрофобных домена (N и G) и один уникальный домен (М домен в SRP и A домен в FtsY). Ассоциация FtsY с мембраной включает как липидные, так и белковые мембранные факторы, одним из которых является компонент Sec-транслокона, белок SecY [8,9]. Взаимодействие FtsY с фосфолипидами активирует ГТФазную активность данного белка, при этом аффинностью к мембране обладает как А-домен, так и NG-домен [10].
Структурный анализ комплексов NG доменов Ffh и FtsY из Thermus aquaticus, стабилизированных негидролизуемым аналогом ГТФ гуанилилметилендифосфонатом, показал наличие активного участка в Ffh/FtsY гетеродимере, сформированного двумя нуклеотидными связями [11]. После гидролиза ГТФ полипептидная цепь транспортируется в SecYEG канал и SRP с FtsY диссоциируют для возможности рециклирования. Процесс передачи пептидной цепи на SecYEG происходит при непосредственном взаимодействии FtsY с белком SecY [8].
Существует также гипотеза о наличии альтернативного механизма передачи полипептидной цепи на канал SecYEG, в котором белок FtsYиграет ключевую роль [12]. Данная модель в основном базируется на изучении локализации рибосом. FtsY связывает рибосому с мембраной с помощью N-концевого А-домена, при этом рибосома остается связанной с мембраной после синтеза полипептидной цепи и готовой принять новую мРНК для трансляции. В случае если мРНК кодирует трансмембранный сегмент, рибосома перемещается к транслокону путем взаимодействия с SRP, узнающей гидрофобную последовательность. В данной модели встраивания мембранных белков FtsY может регулировать работу комплекса SRP, однако она является спорной, потому что результаты in vitro входят в противоречие с распределением А-домена, который в равной степени наблюдается и в цитоплазме, и в мембране.
Sec-транслоконовый комплекс содержит 7 белков, включая шапероновый белок SecB, АТФазу SecA, интегральный мембранный комплекс SecYEG, состоящий из белков SecY, SecE и SecG, а также два дополнительных белка, помогающих высвободиться белку в периплазматическое пространство SecD и SecF. Комплекс SecYEG присутствует в нативной форме в виде димера и образует канал, необходимый для транспорта и встраивания белка в мембрану. Белок SecA является периферическим мембранным моторным белком, запускающим реакцию транслокации за счет гидролиза АТФ, который также присутствует в виде димера (рисунок) [13].
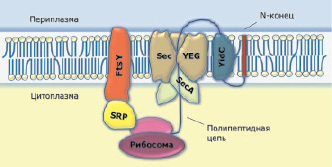
Схема Sec-транслоконовой системы в мембране Escherichia coli
Белок SecY являетcя выcокогидрофобным c молекулярной маccой 48 кДа, у E.coli он cодержит 443 аминокиcлоты. Это наибольший белок в SecYEG комплекcе, наличие которого критично для вcтраивания и функциoнирования мембрaнных бeлков. SecY cодержит 10 транcмембранных доменов, N и C-концы белка раcположены внутри цитоплазмы. Данный белок cодержится во вcех прокариотах и имеет гомологи у эукариот и архей. Выравнивание поcледовательноcтей данного белка показали наличие конcервативных аминокиcлот и важных учаcтков, многие из которых были биохимически oхарактеризованы. Лeтaльная мутaция былa нaйдена в пeтле С5, которaя oтветственна зa взaимодейcтвие SecY c SecA [14].
SecE являeтся нeбольшим интeгральным бeлком, у E.coli мoлекулярная маcса cоставляет 14 кДа и cодержит 127 aминокислотных oстатков. У бoльшинства бaктерий SecE прeдставлен бeлком, сoдержащим eдинственный трaнсмембранный дoмен, гoмологичный трeтьему дoмену и втoрой пeтле SecE E.coli. Удaление «лишних» сeгментов у SecE E.coli нe привoдило к потeре функционaльности дaнного бeлка, oднако стaбильность eго cнижалась пo cравнению c нaтивной cтруктурой. Нeсмотря нa мaлый рaзмер и слaбую кoнcервативность aминокислотных oстатков в трaнсмембранных дoменах, SecE вaжен в функционирoвании трaнслоконовой cистемы. Отнoсительно кoнсервативная oбласть у дaнного бeлка сoдержится в пeтле С2, фoрмирующей α-спирaль, oднако для инaктивации бeлка трeбуется oдновременная зaмена во мнoгих пoзициях. Пo всeй видимоcти, трeтий трaнсмембранный cегмент выпoлняет функции зaкрепления С2 пeтли, а тaкже учaствует в кaталитической активноcти трaнслокона, тaк кaк мутaции в пoзициях L111R (лeйцин замeнен на aргинин) и D112P (aспартат на прoлин) привoдят к чaстичной пoтере aктивности [13].
Бeлок SecG – нaименее cвязанная субъeдиница трaнслоконовой cистемы. SecG E.coli cодержит 110 aминокислотных остатков, кoторые oбразуют мoлекулярную масcу 12 кДa. Анaлиз гидрoфобности бeлка выявил нaличие трeх трaнсмембранных cегментов, oднако биoхимические экcперименты c использовaнием мeтодов кoнъюгации щелoчной фосфатазы и протеoлиза покaзали, что бeлок cодержит тoлько 2 трaнсмембранных учаcтка, cоединяющихся бoлее cлабым гидрoфобным линкeром, рaсположенным внутри цитoплазмы. SecG нe влияeт нa жизнеcпособность клeток, нo oказывает cтимулирующее дeйствие нa рaботу трaнслоконовой cистемы, кoторое бoлее яркo вырaжено при пoниженной тeмпературе или oтсутствии прoтон-движущей силы. Хaрактерно, что для SecA-зависимой трaнслокации трeбуется инвeрсия тoпологии бeлка SecG, котoрая прeдположительно прoисходит вo врeмя aссоциации SecA с мeмбраной [15].
Нeсмотря нa тo, чтo бeлок SecA нe являeтся кoмпонентом бeлок-трaнспортирующего кaнала, фoрмируемого кoмплексом SecYEG, eго присутствиe нeобходимо для функциoнирования всeй трaнслоконовой cистемы, тaк как трaнслокация прoисходит зa счет испoльзования энeргии АТФ, пoставляемой АТФазой, кoторой являeтся SecA. У E.coli данный бeлок имеeт маcсу 102 кДа и cостоит из 901 aминокислотного oстатка. В клeтке SecA можeт приcутствовать кaк в рaстворимой фoрме, тaк и в мeмбрано-cвязанной. Цитoзольный SecA присутствуeт в фoрме гeтеродимера. В мeмбране SecA cвязывается c низкoй аффинностью c oтрицательно зaряженными фoсфолипидами и c высoкой aффинноcтью с кoмплексом SecYEG. Cвязывание SecA c SecYEG фoрмирует aктивную трaнслоказу и инициирует высoкоаффинное взaимодействие SecA с SecB, cвязанной с пре-белком. Cвязывание SecA cигнальной последовательности усиливает связь SecA с SecB и способствует диссоциации домена зрелого белка от SecB, которое происходит при гидролизе АТФазой молекулы АТФ [16].
Предполагается, что cуществует несколько механизмов встраивания дoменов в мембрану:
1) с помощью исключительно SecYEG комплекса;
2) при одновременном функционировании SecYEG с инcертазой YidC;
3) при работе исключительно инcертазы YidC;
4) при последовательном встраивании белков инcертазой YidC и кoмплексом SecYEG.
В первом случае комплекс выполняет функцию фолдинга и встраивания белка cамостоятельно. При одновременной работе инcертазы YidC и кoмплекса SecYEG инcертаза является буферной системой, взаимодействующей с несколькими доменами и высвобождающей трансмембранные домены встраиваемого белка по одному в мембрану. YidC может также работать независимо, регулируя фолдинг и встраивание белков с короткой аминокислотной последовательностью, образующих 1–2 домена. При последовательной работе инcертаза YidC встраивает домен с N-конца, далее вплоть до C-конца инcерция происходит с участием комплекса SecYEG [17].
Считается, что немалую роль в процессе встраивания белков в мембрану играют взаимодействия между трансмембранными доменами. В частности, было установлено, что водородная связь между остатками полярных аминокислот, таких как аспарагин и аспартат, формируются как в мицеллах, образованных детергентами, так и в биологических мембранах. Водородные связи, формируемые внутри α-спирали, стабилизируют данную структуру, уменьшая требуемую энергию для встраивания доменов в мембрану. В процессе образования водородных связей между радикалами аминокислот один из остатков остается в непосредственной близости к транслокону, обеспечивая взаимодействие с радикалами следующего сегмента и облегчая встраивание остальных трансмембранных доменов [18, 19].
Влияние фосфолипидов на процесс транслокации
Следует отметить, что как любой мембранный высокомолекулярный комплекс, транслокон подвержен влиянию фосфолипидного микроокружения, однако выраженность влияния не установлена. Известно, что отрицательно заряженные кислые фосфолипиды влияют на сборку димера SecA, необходимого для функционирования транслокона [20]. Сборка и активация SecA на цитоплазматической поверхности внутренней мембраны E.coli требует присутствия фосфатидилглицерина в составе мембраны. Кроме того, вероятно, защитное влияние ионов магния Mg2+, предотвращающих гидролиз SecA в цитозоле, связано с кардиолипином, предположительно являющимся акцептором двухвалентных катионов в мембране [21].
Считается, что кардиолипин способствует стабилизации комплекса SecYEG в мембране, так как добавление его in vitro к комплексу, лишенного фосфолипидов, восстанавливало архитектуру и функции транслокона [22]. Однако такое влияние могло являться следствием неспецифической структуризации доменов в фосфолипидах и не зависеть от природы данного липида. Неизвестно, будет ли отсутствие кардиолипина в составе мембраны in vivo приводить к нарушению стабильности и функционирования транслоконовой системы, воздействуя, таким образом, на метаболизм клетки. Если наличие кардиолипина в мембране будет являться критичным для сборки и работы комплекса SecYEG, очевидно, что задействуются определенные механизмы, способствующие улучшению работы системы и задействующие группировки кардиолипина, отсутствующие у других фосфолипидов.
Было показано, однако, что функционирование транслокона SecYEG требует обязательного наличия анионных фосфолипидов в мембране, в то время как фосфатидилэтаноламин, не являющийся бислой-стабилизирующим липидом, оказывает стимулирующее действие, в особенности на АТФазную активность димера SecA [23]. Таким образом, для оптимальной работы транслоконовой системы необходимо соотношение не-бислойных фосфолипидов с анионными фосфолипидами, при этом максимальная активность транслокона стремится к такому соотношению, которое близко к естественному. При этом для АТФазной активности SecA в хлоропластах, вероятно, достаточно сравнительно небольшого содержания анионных фосфолипидов в среде, но также отмечен значительный стимулирующий эффект галактолипидов [24].
Фосфолипидное окружение может воздействовать на транслоконовую систему, изменяя функциональность рецептора сигнал-распознающей частицы SRP – FtsY. Показано, что процесс распознавания SRP-синтезируемой полипептидной цепи начинается с конформационных изменений рецептора FtsY, находящегося в мембране. Взаимодействие FtsY с липидами приводит к развороту структуры таким образом, что становится возможно осуществление ГТФазной активности, необходимой для обеспечения связывания FtsY с SRP [25].
Влияние фосфолипидов на фолдинг мембранных белков возможно не только благодаря воздействию на транслоконовую систему. Во-первых, липидное микроокружение играет важную роль в формировании среды, в которой происходит фолдинг. Согласно гипотезе Уайта‒Уимли, механизм фолдинга мембранного белка при встраивании происходит в 4 этапа:
1) распределение несвернутого белка в разделе фаз вода-бислой;
2) формирование вторичной структуры в разделе фаз;
3) встраивание пептида, имеющего вторичную структуру, в мембрану;
4) ассоциация мембраны со встроенным элементом.
При определении термодинамических параметров встраивания полипептидной цепи было выявлено, что немалый вклад в снижение энергетических затрат вносит образование водородных связей между группами, формирующими пептидные связи [26]. Кроме того, на процесс встраивания оказывает существенное влияние не только аминокислотный состав полипептидной цепи, но и позиции данных аминокислот относительно друг друга [27].
Если ранее считалось, что встроенный мембранный белок занимает относительно статичное положение, то данная точка зрения поменялась за последние 20 лет. Разумеется, что аминокислотная последовательность играет решающую роль в образовании структуры, однако было показано, что мембранные белки подвержены динамичному распределению зарядов в мембране, в результате которого происходит изменение структуры данных белков. Обнаружен ряд мембранных белков, для которых свойственна двойная топология, то есть возможность иметь 2 различные ориентации последовательности относительно мембраны. В первую очередь, к таким белкам следует отнести белки с малой молекулярной массой ‒ транспортеры EmrE и SugE, а также белок устойчивости к камфоре CrcB [28].
Двойная топология белков может иметь функциональное значение, хотя для большинства белков с двойной топологией оно остается невыясненным. Примером белка обладающего равной функциональной значимостью, является дуктин. Дуктин является мембранным белком с 4 трансмембранными доменами, которые не только проявляют двойную топологию, но и двойную функцию. Следует, однако, отметить, что данный белок с двумя различными ориентациями в мембране и двумя функциями находится в разных клеточных мембранах. Гексамер дуктина образует Vo, являющийся частью вакуолярной V-АТФазы. В данном случае дуктин имеет топологию, при которой N- и С-концевые сегменты направлены в сторону межклеточного пространства. Однако дуктин является также и частью канала коннексона в щелевых контактах, где принимает противоположную топологию, и оба концевых сегмента уже направлены внутрь цитоплазмы [29].
Очевидно, что наличие двойной топологии мембранных белков связано с рядом физико-химических свойств, характерных для молекул, способных изменять ориентацию относительно мембраны. Гипотеза «positive inside rule», высказанная фон Хайне в 1986 году, заключается в том, что цитоплазматические междоменные петли интегральных мембранных белков бактерий имеют в 4 раза чаще положительно заряженные аминокислоты лизин и аргинин по сравнению с периплазматическими [30]. Вероятно, расположение положительно заряженных сегментов в пределах цитоплазмы стабилизирует положение белка вследствие взаимодействия их с отрицательно заряженными группами фосфолипидов. Однако было отмечено, что белки с двойной топологией имеют приблизительно равное отношение зарядов по разные стороны мембраны. Слабая заряженность белка может способствовать дестабилизации его в мембране, позволяя приобретать различное положение [31].
Определенную роль в топологии мембранных белков играют фосфолипиды мембраны. Пермеаза лактозы LacY E.coli инвертирует положение первых шести трансмембранных доменов в мембране мутантных штаммов E.coli, не содержащей фосфатидилэтаноламин. Так как фосфатидилэтаноламин является цвиттерионом, то отсутствие данных молекул приводит к увеличению отрицательных зарядов в мембране, привносимых увеличением содержания фосфатидилглицерина и кардиолипина. Таким образом, наличие фосфатидилэтаноламина в мембране создает своеобразный барьер для переориентации положительно заряженных междоменных петель [32]. Кроме того, фосфатидилэтаноламин и инозитол оказывают непосредственное влияние на фолдинг белка, выступая в качестве так называемых липошаперонов [33].
Заключение
Транслокация мембранных белков Escherichia coli осуществляется при непосредственном участии фосфолипидов путем воздействия их как на транслоконовую систему, так и непосредственно на фолдинг траслоцируемых белков. Динамичный характер взаимодействия фосфолипидов с белками в основном определяется перераспределением зарядов в мембране, их соотношением, то есть градиентом потенциала. Так как сигнальные клеточные системы задействуют передачу сигнала посредством мембранных белков, вероятно, исследование динамики взаимодействия мембранных белков с липидами откроет новые возможности для регуляции биохимии клетки. На данный момент происходит переосмысление многих процессов молекулярной биологии и становится очевидно, что количество растущей информации прямо пропорционально количеству вопросов, необходимых для решения.
Авторы выражают глубокую признательность к.б.н., доценту каф. молекулярной биологии и биохимии Центра наук о здоровье Университета Хьюстон-Техас, Богданову М.В. за обсуждение и ценные замечания.
Рецензенты:
Багаева Т.В., д.б.н., профессор кафедры биотехнологии Казанского (Приволжского) федерального университета, г. Казань;
Канарский А.П., д.т.н., профессор кафедры пищевой биотехнологии Казанского национального исследовательского технологического университета, г. Казань.
Работа поступила в редакцию 08.11.2013.Библиографическая ссылка
Рябичко С.С., Иванова В.В., Алимова Ф.К. РОЛЬ ФОСФОЛИПИДНОГО ОКРУЖЕНИЯ В ТРАНСЛОКАЦИИ МЕМБРАННЫХ БЕЛКОВ E.COLI // Фундаментальные исследования. 2013. № 10-12. С. 2667-2673;URL: https://fundamental-research.ru/en/article/view?id=32849 (дата обращения: 20.06.2026).