


Scientific journal
Fundamental research
ISSN 1812-7339
"Перечень" ВАК
ИФ РИНЦ = 1,798
DYNAMICS OF ACCUMULATION OF BIOCHEMICAL COMPOUNDS ANTIOXIDANT ACTION IN DIFFERENT ORGANS OF BUCKWHEAT DURING ONTOGENY

За последние десятилетия область научных знаний расширяет свои границы в направлении углубленного изучения фундаментальных процессов, происходящих в живых организмах: растениях, животных и человеческом организме. Все жизненные процессы, связанные с потреблением кислорода, в той или иной степени способствуют образованию свободных радикалов. Перекисное окисление липидов или же цепное окисление ненасыщенных жирных кислот с участием молекулярного кислорода один из основных процессов образования свободных радикалов. Известно, что умеренная активация перекисного окисления липидов в ответ на действие поражающего фактора является одним из адаптационных механизмов и выполняет регуляторную функцию. Она направлена на увеличение проницаемости клеточной мембраны, облегчение работы мембранных белков, но, превышая определенные пределы, увеличение количества свободных радикалов, приобретает саморазрушающее действие, которое проявляется денатурацией и инактивацией белка, делипидизацией мембраны, нарушением деления и роста клетки [5]. Ход нормального развития физиологических и биохимических процессов в жизнедеятельности клетки контролируется антиоксидантной системой. Вещества, способные преобразовывать свободные радикалы из активной в неактивную, инертную форму, называют антиоксидантами. К антиоксидантам относят вещества различных химических классов, как высокомолекулярные соединения белковой природы – ферменты, так и низкомолекулярные соединения. Разные антиоксиданты по-разному работают в организме: возможно непосредственное связывание со свободными радикалами и опосредованное увеличение устойчивости мембранной структуры клетки. Антиоксиданты могут синтезироваться в организме и поступать извне. В идеале объединение системы перекисного окисления липидов и антиоксидантной системы должно находиться в равновесном движении всех физиолого-биохимических процессов, таким образом, у живого организма повышается устойчивость к воздействию стресс-факторов и формируется адаптация в меняющихся условиях окружающей среды [6]. Среди соединений с антиоксидантной активностью выделяют группу природных фенольных соединений, в число которых входят флавоноиды.
Флавоноидами являются производные бензо-γ-пирона, в основе которых лежит скелет, состоящий из двух бензольных колец, соединенных между собой трехуглеродной цепочкой, общей формой С6-С3-С6.
Флавоноиды широко распространены в растительном мире. Они обнаружены почти во всех высших растениях. Особенно богаты флавоноидами высшие растения, к которым относятся софора японская, рута и гречиха. Целью наших исследований являлось изучение сортовых особенностей гречихи посевной по накоплению комплекса флавоноидов и определение механизмов накопления флавоноидов.
Материалы и методы исследования
Полевой опыт проводится на полях ГНУ ВНИИ ЗБК Орловской области (п. Стрелецкий). Площадь опытной делянки 1 кв. м. Повторность опытов – четырехкратная. Посев семян гречихи проводится рядовым способом на глубину 3 см при прогревании почвы до 15–18 °С и оптимальной влажности 60–70 %. Норма высева семян из расчета 1,5 млн шт./га. Уход за посевами проводится в соответствии с технологией возделывания гречихи посевной в Центрально-черноземном регионе. Лабораторные опыты проводили в 3-кратной биологической повторности, аналитическое определение для каждой пробы – в трех повторностях.
Для определения активности супероксиддисмутазы использовали модифицированную методику с использованием прибора – фотореактора (Giannopolities C.N., 1977). Определение МДА проводили по реакции с тиобарбитуровой кислотой по образованию окрашенного триметинового комплекса, с максимумом поглощения при 532 нм. Молярный коэффициент экстинкции этого комплекса: ε = 1,56∙105 см–1∙М–1. Определение рутина, ПФС и флавонолов в растениях гречихи посевной выполняли по Ермакову [2].
Достоверность экспериментальных данных оценивали методами математической статистики с привлечением современных программных средств Excel 7 для Windows ХР.
Результаты исследования и их обсуждения
Накоплению флавоноидов способствуют умеренная влажность и температура, высота над уровнем моря и почва, богатая азотом, калием, фосфором и др. В южных и высокогорных районах под влиянием света и на почвах, богатых микроэлементами, в растениях содержание флавоноидов выше [3, 4]. Флавоноиды обнаружены в различных частях и органах растения гречихи посевной. Больше они накапливаются в надземной части растения. Анализ соотношения рутина, флавонолов и полифенольных соединений (ПФС) в плодах представлен на рис. 1. На диаграмме видна высокая степень отличия содержания веществ по сортам.
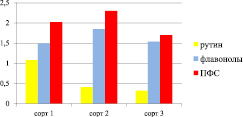
Рис. 1. Содержание рутина, флавонолов и полифенольных соединений в плодах разных сортов гречихи, мг/г
Фенольные соединения принимают участие в окислительно-восстановительных процессах и в процессе фотосинтеза. Являясь растительными пигментами, флавоноиды (антоцианы) придают яркую окраску цветкам, чем привлекают насекомых и тем самым способствуют опылению и размножению растений. В растениях флавоноиды содержатся чаще всего в виде гликозидов, которые растворены в клеточном соке, сосредоточены в вакуолях и хлоропластах. Наиболее богаты ими молодые цветки, незрелые плоды.
Максимальное накопление флавоноидов в надземной части гречихи – в период бутонизации и цветения, затем содержание флавоноидов снижается. Содержание флавоноидов в генотипах гречихи различно в среднем 0,5–5 % [1].
В настоящее время практически отсутствуют публикации о выяснении вклада различных групп низкомолекулярных соединений в функционирование клеточной антиоксидантной системы. Флавоноиды в нормальных условиях участвуют в поглощении световой энергии, а при стрессе являются частью фотозащитной и антиоксидантной систем.
Сложность изучения данной проблемы заключается в том, что усиление биосинтеза одного из низкомолекулярных соединений влечет за собой изменения в метаболизме в целом и перераспределение потребляемых субстратов при изменении энергетических затрат. В настоящее время мало изучено функционирование антиоксидантных ферментов и флавоноидов. Существуют два пути адаптации растений: усиление активности антиоксидантных ферментов и второй путь – накопление низкомолекулярных метаболитов.
Поскольку флавоноиды локализуются в эпидермальных клетках, исследования динамики их содержания проводили в листьях и в корнях гречихи посевной – органах, отличающихся накоплением рутина.
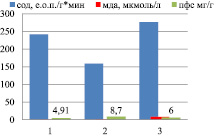
Сравнительный анализ в вегетативный и генеративный периоды по уровню МДА, СОД и ПФС в корнях показал снижение накопления полифенольных соединений при снижении уровня МДА и повышении активности СОД (рис. 2). Такая тенденция прослеживается как в индетерминантных, так и детерминантных сортах гречихи посевной.
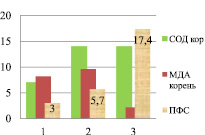
Индетерминантные сорта: 1 – Баллада, 2 – Молва, 3 – Башкирская красностебельная
Детерминантные сорта: 1 – Дизайн, 2 – Дикуль, 3 – Дождик, 4 – Деметра, 5 – Диалог, 6 – Темп, 7 – Уша
Рис. 2. Уровень МДА, СОД и ПФС в корнях районированных сортов гречихи посевной (вегетативный период)
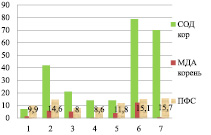
В листьях накопление ПФС происходит при повышении активности СОД и повышении уровня МДА от вегетативного к генеративному периоду (рис. 3)
Индетерминантные сорта: 1 – Баллада, 2 – Молва, 3 – Башкирская красностебельная
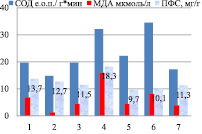
Детерминантные сорта: 1 – Дизайн, 2 – Дикуль, 3 – Дождик, 4 – Деметра, 5 – Диалог, 6 – Темп, 7 – Уша
Рис. 3. Уровень МДА, СОД и ПФС в листьях районированных сортов гречихи посевной (вегетативный период – фаза всходы)
Вегетативный период онтогенеза гречихи посевной, отличающийся низким накоплением количества флавоноидов, он характеризуется низкой активностью СОД и высоким уровнем МДА: в индетерминантных сортах СОД (от 12 до 17 е.о.п./г∙мин в листьях и от 7 до 14 е.о.п./г∙мин в корнях) (р < 0,05) – МДА (от 3 до 5 мкмоль/л в листьях и 8,1 и 9,8 мкмоль/л в корнях) (р < 0,01); в детерминантных сортах СОД (от 18 до 34 е.о.п./г∙мин в листьях и от 8 до 77 е.о.п./г∙мин в корнях) (р < 0,01) и МДА – (от 3 до 18 в листьях и от 3 до 15 мкмоль/л в корнях) (р < 0,05), (рис. 3).
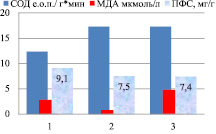
Индетерминантные сорта: 1 – Баллада, 2 – Молва, 3 – Башкирская красностебельная
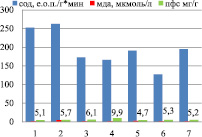
Детерминантные сорта: 1 – Дизайн, 2 – Дикуль, 3 – Дождик, 4 – Деметра, 5 – Диалог, 6 – Темп, 7 – Уша
Рис. 4. Уровень МДА, СОД и ПФС в корнях районированных сортов гречихи посевной (генеративный период – фаза цветение)
Сорта Темп и Уша выделяются по накоплению ПФС в корнях (15,1 и 15,7 мг/г), имеют повышенную активность СОД.
Генеративный период онтогенеза с интенсивным накоплением флавоноидов характеризуется резким скачком активности СОД (от 180 до 260 е.о.п./г∙мин) (р < 0,05) и снижением уровня МДА в корнях (рис. 4), но повышением уровня МДА в листьях (рис. 5). Индетерминантный сорт Башкирская красностебельная отличается высоким уровнем накопления ПФС в корнях (17,4 мг/г), так как при низкой активности СОД наблюдается высокий уровень МДА, что способствует повышенному уровню накопления флавоноидов.
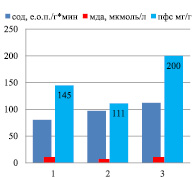
Индетерминантные сорта: 1 – Баллада, 2 – Молва, 3 – Башкирская красностебельная
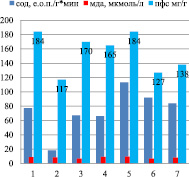
Детерминантные сорта: 1 – Дизайн, 2 – Дикуль, 3 – Дождик, 4 – Деметра, 5 – Диалог, 6 – Темп, 7 – Уша
Рис. 5. Уровень МДА, СОД и ПФС в листьях районированных сортов гречихи посевной (генеративный период – фаза цветение)
Вывод
Процесс онтогенеза гречихи посевной районированных сортов Орловской области характеризуется сменой работы ферментативных систем с изменением активности в зависимости от биологического возраста и локализации по органам растений гречихи посевной. Динамика флавоноидов в ходе онтогенеза находится в зависимости от работы антиоксидантной системы гречихи посевной, и накопление флавоноидов связано с увеличением интенсивности работы высокомолекулярных и низкомолекулярных компонентов антиоксидантной системы растения.
Рецензенты:
Ярован Н.И., д.б.н., профессор, заведующая кафедрой химии, ФГБОУ ВПО «Орловский государственный аграрный университет», г. Орел;
Козлов А.С., д.б.н., профессор, заведующий кафедрой зоогигиены и кормления сельскохозяйственных животных, ФГБОУ ВПО «Орловский государственный аграрный университет», г. Орел.
Работа поступила в редакцию 15.08.2013.
Библиографическая ссылка
Полехина Н.Н., Павловская Н.Е. ДИНАМИКА НАКОПЛЕНИЯ БИОХИМИЧЕСКИХ СОЕДИНЕНИЙ АНТИОКСИДАНТНОГО ДЕЙСТВИЯ В РАЗНЫХ ОРГАНАХ ГРЕЧИХИ В ПРОЦЕССЕ ОНТОГЕНЕЗА // Фундаментальные исследования. 2013. № 10-2. С. 357-361;URL: https://fundamental-research.ru/en/article/view?id=32283 (дата обращения: 28.06.2026).