


Scientific journal
Fundamental research
ISSN 1812-7339
"Перечень" ВАК
ИФ РИНЦ = 1,798
SALT-TOLERANCE PARAMETERS OF SOME FOREST-STEPPE MEDICINAL PLANTS

В системе экологической специализации видов растений мировой флоры особое место занимают солеустойчивые растения – группа физиологически и биохимически специализированных видов, способных нормально функционировать и продуцировать в условиях засоленной среды. Новая стратегия растениеводства рассматривает солеустойчивые лекарственные растения как ресурс для возделывания на засоленных почвах, где традиционные культуры не могут произрастать. Селекционный потенциал мировой солеустойчивой флоры определяется не только числом видов, населяющих регион или экологическое местообитание, но и в не меньшей мере совокупностью внутривидовых эколого-физиологических и биохимических особенностей растений. Генетические ресурсы галофитов России пока еще слабо изучены и оценены [8].
В основе солеустойчивости растений находятся эффективные механизмы солевого обмена. Для галофитов характерна аккумулирующая способность в отношении водорастворимых ионов, которые понижают водный потенциал клеток, облегчая поступление воды. Перераспределение солей по органам растений осуществляется с затратой метаболической энергии, при участии переносчиков, встроенных в клеточные мембраны. Покрытосеменные галофиты накапливают ионы в листьях. Среди гликофитов наиболее устойчивы к избытку солей формы, в корнях которых идет аккумуляция ионов [1, 3, 4, 6, 7, 9, 10].
Возможность культивации лекарственных трав на засоленных почвах опирается на эффективность адаптивных механизмов растений. Солеустойчивость внутривидовых экотипов растений можно установить на территориях с природными популяциями. В лесостепном заказнике «Троицкий», расположенном на юге Челябинской области, 43,7 % площади занимают луговые солонцы и около 5 % – солончаки и солончаковые почвы, на которых обитают некоторые виды лекарственных растений.
Цель исследований – оценить адаптивный потенциал к засолению термопсиса ланцетовидного (Thermopsis lanceolata, R. Br), кровохлебки лекарственной (Sanguisorba officinalis L.), солодки Коржинского (Glycyrrhiza korshinskyi, Grig.), обитающих на луговых почвах в зауральской лесостепи [5].
Материалы и методы исследований
На территории заказника «Троицкий» в популяциях лекарственных трав в фазу вегетации – начала цветения отбирали почвенные и растительные пробы. Почвенные пробы взяты с глубины 0–20 см. В водной вытяжке из почв определили общую щелочность и количество Cl- – титриметрическим методом, Ca2+ и Mg2+ – комплексометрическим методом, Na+ – методом пламенной фотометрии, SO42– установлены по разности сумм катионов и анионов; сумма солей – расчетная. Для каждого вида исследованы почвы с 16-ти мест обитания.
В водной вытяжке из листьев и подземных органов растений определяли количество Сl– – меркурометрическим методом, Na+ – методом пламенной фотометрии. Всего проанализировано от 16 до 24 проб листьев и корневищ каждого вида растений.
Результаты исследований и их обсуждение
На территории заказника популяции исследуемых лекарственных трав обитают на луговых почвах с близким залеганием грунтовых вод. В местах произрастания лекарственных растений слой почвы 0–20 см характеризовался низким содержанием солей. Химизм засоления почвы в соответствии с составом ионов был гидрокарбонатно-сульфатно-хлоридным натриевым под термопсисом, смешанным, при равном участии анионов, кальциево-натриевым – под кровохлебкой, гидрокарбонатно-хлоридным кальциево-натриевым – под солодкой (рисунок).
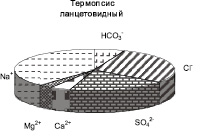
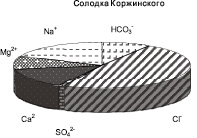
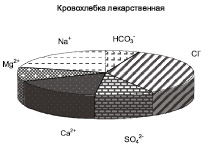
Соотношение ионов в водной вытяжке из почв (слой 0–20 см) в местах произрастания лекарственных трав
Содержание водорастворимого Na+ в почвах колебалось в диапазоне от 0,25 до 5,35 мг∙экв/100 г почвы. Термопсис встречался на почвах c более высоким уровнем содержания натрия, там, где популяции двух других видов не произрастали (табл. 1). Солодка и кровохлебка произрастали при одинаковом уровне содержания Na+ в слое почвы 0–20 см, однако популяции последней обнаружены при более высоком количестве натрия в почве.
Содержание водорастворимых Cl- в почвенных слоях 0-20 см варьировалось от 0,77 до 5,67 мг-экв/100 г почвы. Термопсис произрастал при большем количестве Cl- в почве, популяции солодки и кровохлебки встречались при одинаковом засолении хлоридами (табл. 1).
Общая щелочность водной вытяжки из корнеобитаемых слоев почв варьировалась в пределах от 0,09 до 1,27 мг-экв/100 г почвы. Под термопсисом почва характеризовалась повышенным уровнем щелочности. Доверительные интервалы общей щелочности под популяциями солодки и кровохлебки приблизительно совпадали (табл. 1).
Таблица 1
Содержание водорастворимых ионов в корнеобитаемом слое почв в местах обитания лекарственных растений, мг-экв/100 г
|
Показатель |
Na+ |
Cl– |
HCO3– |
Сумма солей, % |
|
Термопсис ланцетовидный |
||||
|
Среднее значение |
3,30 |
2,52 |
0,52 |
0,26 |
|
95 % -й доверительный интервал |
2,69–3,91 |
2,03–3,02 |
0,40–0,63 |
0,22–0,31 |
|
Кровохлебка лекарственная |
||||
|
Среднее значение |
0,82 |
1,41 |
0,23 |
0,13 |
|
95 % -й доверительный интервал |
0,56–1,09 |
1,29–1, 52 |
0,17–0,30 |
0,11–0,14 |
|
Солодка Коржинского |
||||
|
Среднее значение |
0,54 |
1,37 |
0,27 |
0,09 |
|
95 % -й доверительный интервал |
0,43–0,65 |
1,22–1,53 |
0,19–0,34 |
0,08–0,10 |
Содержание водорастворимых SO42– в почвах под популяциями лекарственных растений варьировалось от 0,07 до 3,35 мг∙экв/100 г почвы, достигало наибольших величин в местах произрастания термопсиса ланцетовидного.
Сумма водорастворимых солей в слое 0–20 см варьировала от 0,09 до 0,36 % (табл. 1). Термопсис произрастал при наиболее высоком засолении, что указывает на его относительно повышенную солеустойчивость. Несколько меньше сумма солей в луговых почвах под кровохлебкой. Солодка произрастала при минимальном количестве легкорастворимых солей и узком интервале изменчивости их содержания в почве.
Таким образом, установлено, что на территории заказника лекарственные растения произрастали в условиях низкого засоления при смешанном химизме (с участием HCO3–, SO42–, Cl–), термопсис ланцетовидный – при повышенном содержании хлоридов, сульфатов натрия и общей щелочности почвы по сравнению с двумя другими видами трав. Кровохлебка лекарственная и солодка Коржинского обитали при относительно пониженном засолении, при этом кровохлебка произрастала при несколько более высокой концентрации солей натрия в почве.
В условиях засоленной почвенной среды в растениях накапливается значительное количество ионов Na+ и Сl–. Определенное количество Na+ необходимо для формирования сосудистой и механической систем растений. Натрий наряду с другими ионами играет осмотическую и ионбалансирующую роль, выполняет более специфическую функцию в конформационных изменениях ферментов и при катализе. Однако при засолении почвы может идти избыточное накопление Na+. Происходит чрезмерная гидратация коллоидов пластид и протоплазмы ассимилирующих клеток, что приводит к снижению интенсивности фотосинтеза. С увеличением содержания натрия образование биоколлоидов плазмы сильно увеличивается. Повышение концентрации Na+ в цитоплазме нарушает ход метаболических процессов из-за вызываемого им ионного дисбаланса и токсичности натрия. Ионы Сl– в растениях находятся преимущественно в ионной форме, участвуют в осморегуляции и ионном гомеостазе [1, 2, 6].
У термопсиса ланцетовидного количество Na+ варьировалось в листьях от 302 до 1011 мг/100 г с.м., в подземных органах от 385 до 794 мг/100 г с.м., достоверных различий между органами по содержанию иона не установлено (табл. 2).
Содержание Cl– в органах термопсиса колебалось от 1093 до 3642 мг/100 г с.м. и заметно превышало накопление натрия (табл. 2). В корнях содержится в 2–3 раза меньше свободных ионов хлора, чем в надземных органах. Достоверность различий между надземными органами и корнями подтверждает активный механизм поглощения из почвы и транспорта ионов хлора в листья. Широкий интервал накопления Cl- в листьях, вероятно, указывает на способность термопсиса произрастать в достаточно большом диапазоне засоления почвы этим ионом. Явление дифференцированного поглощения Cl- с преимущественным накоплением в надземных органах выражено у солеустойчивых растений лесостепного заказника, при этом они значительно больше аккумулируют Cl-, чем Na+ (Еремченко и др., 2004). Следовательно, уровень накопления Cl–, их дифференциация в органах отражает определенную солеустойчивость термопсиса ланцетовидного.
Содержание Na+ в органах кровохлебки в среднем невысокое и варьировалось от 47 до 493 мг/100 г с.м., листья и корневища существенно не отличались по этому иону (табл. 2). Количество Cl- в органах кровохлебки заметно выше, чем натрия, и колебалось в пределах от 1134 до 1765 мг/100 г с.м. (табл. 2). Сравнительный анализ показывает, что уровень аккумуляции ионов хлора сопоставим с солеустойчивыми растениями, произрастающими на солонцах заказника, однако отсутствует избирательная аккумуляция в листьях по сравнению с корневищами.
Таблица 2
Накопление Na+ и Cl– в органах лекарственных растений, мг/100 г сухой массы
|
Органы растений |
Na+ |
Cl– |
||
|
среднее |
95 %-й доверительный интервал |
среднее |
95 %-й доверительный интервал |
|
|
Термопсис ланцетовидный |
||||
|
Листья |
672 |
549–796 |
2344* |
1992–2696 |
|
Подземные |
629 |
561–698 |
1094 |
938–1251 |
|
Кровохлебка лекарственная |
||||
|
Листья |
164 |
97–232 |
1399 |
1329–1469 |
|
Подземные |
138 |
100–177 |
1210 |
1111–1309 |
|
Солодка Коржинского |
||||
|
Листья |
131* |
104–157 |
1634 |
1469–1799 |
|
Подземные |
166 |
139–193 |
1549 |
1307–1791 |
Примечание. * – достоверные различия по содержанию ионов в листьях и корневищах.
Содержание Na+ в органах солодки колеблется в пределах от 65 до 259 мг/100 г с.м. Количество Na+ в корневищах солодки достоверно повышено по сравнению с листьями. Содержание Cl– в органах солодки выше по сравнению с Na+ и изменялось в значительном интервале от 625 до 2804 мг/100 г с.м., на этом фоне достоверной разницы между органами по количеству ионов не установлено (табл. 2)
Выводы
1. Популяции лекарственных растений показали определенную солеустойчивость, произрастая в условиях засоления корнеобитаемого слоя луговых почв зауральской лесостепи. Термопсис ланцетовидный встречался при наибольшем содержании хлоридов, сульфатов натрия и общей щелочности почв. Кровохлебка лекарственная обитала в местах несколько большего засоления почв по сравнению с солодкой Коржинского.
2. Солеустойчивость термопсиса ланцетовидного обусловлена значительным накоплением ионов Na+ и Cl– в органах, селективным поглощением Cl– по сравнению с Na+, избирательной аккумуляцией Cl– в листьях по сравнению с корневищем.
2. У кровохлебки лекарственной установлено относительно пониженное количество ионов Na+, селективное накопление Cl- по сравнению с Na+, повышенная аккумуляция Cl– в листьях по сравнению корневищами.
3. Наименее солеустойчивая солодка Коржинского характеризовалась селективным накоплением Cl– по сравнению с Na+, повышенным накоплением Na+ в корневище относительно листьев.
Рецензенты:
Боронникова С.В., д.б.н., заведующая кафедрой ботаники и генетики растений, Пермский государственный национальный исследовательский университет, Министерство образования и науки РФ, г. Пермь;
Ширшев С.В., д.м.н., профессор, заведующий лабораторией иммунорегуляции, Институт экологии и генетики микроорганизмов УрО РАН, г. Пермь.
Работа поступила в редакцию 01.08.2013.
Библиографическая ссылка
Еремченко О.З., Кусакина М.Г., Чудинова Л.А. ПОКАЗАТЕЛИ СОЛЕУСТОЙЧИВОСТИ НЕКОТОРЫХ ЛЕСОСТЕПНЫХ ЛЕКАРСТВЕННЫХ РАСТЕНИЙ // Фундаментальные исследования. 2013. № 10-1. С. 105-109;URL: https://fundamental-research.ru/en/article/view?id=32225 (дата обращения: 04.07.2026).