


Scientific journal
Fundamental research
ISSN 1812-7339
"Перечень" ВАК
ИФ РИНЦ = 1,798
MOLECULAR IDENTIFICATION OF SPRING WHEAT GENOTYPES BY ALLELIC VARIANTS OF THE HMW GLUTENIN SUBUNITS

Идентификация генотипов пшеницы по аллельным вариантам HMW субъединиц глютенинов является важным звеном в маркер-вспомогательной селекции сортов с высокими мукомольно-хлебопекарными качествами зерна [1, 2, 3, 4, 5].
Наиболее высокоточными подходами к оценке аллельного полиморфизма HMW субъединиц глютенинов служат способы идентификации на основе молекулярно-генетических методов исследования [1, 2, 3, 4, 5].
Так, одним из подходов к идентификации аллельных вариантов HMW-GS пшеницы является способ проведения ПЦР с праймерами: UMN19F + UMN19R (Ax1/Axnull- и Ax2*-аллели), UMN25F + UMN25R (Dx2- и Dx5-аллели) и UMN26F + UMN26R (Dy10- и Dy12-аллели) с последующей детекцией результатов реакции преимущественно методами капиллярного или вертикального гель-электрофореза в ПААГ [4].
Цель настоящей работы – молекулярная идентификация генотипов яровой пшеницы селекции ТатНИИСХ по аллельным вариантам HMW субъединиц глютенинов и апробация разработанного нами способа проведения ПЦР-ПДРФ-генотипирования с электрофорезной детекцией в агарозном геле.
Материалы и методы исследования
Молекулярно-генетическая оценка 70 образцов яровой пшеницы преимущественно селекции ТатНИИСХ на предмет идентификации генотипов по аллельным вариантам HMW субъединиц глютенинов (HMW-GS) проведена методами ПЦР- и ПЦР-ПДРФ-анализа на основе общеизвестного [4] и разработанного нами способов генотипирования.
Экстракция геномной ДНК из зерновок яровой пшеницы молочно-восковой спелости урожая 2012 г. осуществлена коммерческим набором «ДНК-сорб С» («ЦНИИ эпидемиологии», Россия).
Амплификация геномной ДНК проведена на термоциклере «PTC-200» (MJ Research) с использованием праймеров [4, 5], перечень которых представлен в табл. 1.
Таблица 1
Условия проведения ПЦР- и ПЦР-ПДРФ-анализа для идентификации аллельных вариантов HMW субъединиц глютенинов пшеницы
|
Праймеры |
Последовательности праймеров (5/-3/) |
Локус (аллели) |
Режим амплификации |
ПДРФ-анализ |
|
UMN19F |
CGAGACAATATGAGCAGCAAG |
Glu-A1 (Ax1/Axnull, Ax2*) |
×1:94 °С – 4 мин ×40:94 °С – 30 с, 60 °С – 30 с, 72 °С – 30 с ×1:72 °С – 5 м |
HaeIII 37 °C – 2 часа |
|
UMN19R |
CTGCCATGGAGAAGTTGGA |
|||
|
UMN25F |
GGGACAATACGAGCAGCAAA |
Glu-D1 (Dx2, Dx5) |
×1:94 °С – 4 мин ×40:94 °С – 30 с, 60 °С – 30 с, 72 °С – 30 с ×1:72 °С – 5 мин |
HaeIII 37 °C – 2 часа |
|
UMN25R |
CTTGTTCCGGTTGTTGCCA |
|||
|
UMN26F |
CGCAAGACAATATGAGCAAACT |
Glu-D1 (Dy10, Dy12) |
×1:94 °С – 4 мин ×40:94 °С – 30 с, 60 °С – 30 с, 72 °С – 30 с ×1:72 °С – 7 мин |
HaeIII 37 °C – 2 часа |
|
UMN26R |
TTGCCTTTGTCCTGTGTGC |
|||
|
Axnull-F |
ACGTTCCCCTACAGGTACTA |
Glu-A1 (Axnull) |
×1:94 °С – 4 мин ×40:94 °С – 1 мин, 58 °С – 1 мин, 72 °С – 1 мин ×1:72 °С – 7 мин |
|
|
Axnull-R |
TATCACTGGCTAGCCGACAA |
Детекция результатов ПЦР- и ПЦР-ПДРФ-анализа выполнена методом горизонтального электрофореза в 3 % агарозном геле в буфере ТBE (рН 8,0), содержащем этидий бромид с последующей визуализацией результатов в ультрафиолетовом трансиллюминаторе (λ = 310 нм).
Размеры фрагментов ДНК оценены по подвижности в сравнении со стандартными ДНК маркерами. В работе использованы реактивы для молекулярно-биологических исследований производства ООО «СибЭнзим» (Россия).
Выравнивание частичных нуклеотидных последовательностей аллелей HMW субъединиц глютенинов: CLUSTAL W (v. 1.83). ПЦР-ПДРФ-моделирование: NEBcutter v.2.0.
Результаты исследования и их обсуждение
По результатам практических исследований, направленных на апробацию предложенного способа проведения ПЦР-ПДРФ, нами получен обеспечиваемый заявленным способом технический результат, выраженный в эффективной идентификации аллельных вариантов HMW-GS, ввиду корректной интерпретации при электрофорезной детекции в агарозном геле генерируемых ПЦР-ПДРФ-фрагментов (рис. 2, 4, 6), сопоставимых с расчетными данными (рис. 1, 3, 5).
Отличительным признаком предложенного способа генотипирования от прототипа [4] является дополнительное введение этапа ПДРФ-анализа с эндонуклеазным расщеплением ампликонов рестриктазой HaeIII с последующей детекцией результатов анализа методом горизонтального электрофореза в агарозном геле.
В результате молекулярно-генетической оценки на предмет идентификации генотипов по аллельным вариантам Glu-A1-локуса HMW субъединиц глютенинов установлено, что из 70 происследованных образцов яровой пшеницы 13 растений (18,6 %) имели субъединицу Ax1, кодируемую аллельным вариантом Glu-A1a (Ax1-аллель), а 57 образцов (81,4 %) – субъединицу Ax2*, кодируемую аллельным вариантом Glu-A1b (Ax2*-аллель) (табл. 2).
При оценке этих же образцов яровой пшеницы по Glu-D1-локусу HMW-GS выяснено, что 44 растения (62,9 %) характеризовались наличием комбинации субъединиц Dx5 и Dy10 (5 + 10), кодируемой аллельным вариантом Glu-D1d (Dx5- и Dy10-аллели), а 26 образцов (37,1 %) имели комбинацию субъединиц Dx2 и Dy12 (2 + 12), кодируемую аллельным вариантом GluD1a (Dx2- и Dy12-аллели) (см. табл. 2).
Распределение же исследуемых генотипов по совокупной комбинации Glu-A1-/Glu-D1-локусов HMW субъединиц глютенинов было следующим: Ax1/5 + 10 = 12 (17,1 %); Ax1/2 + 12 = 1 (1,4 %); Ax2*/5 + 10 = 32 (45,7 %); Ax2*/2 + 12 = 25 (35,7 %) образцов яровой пшеницы; с преобладанием желаемой для селекции на мукомольно-хлебопекарные качества зерна комбинацией субъединиц Ax2*/5 + 10.
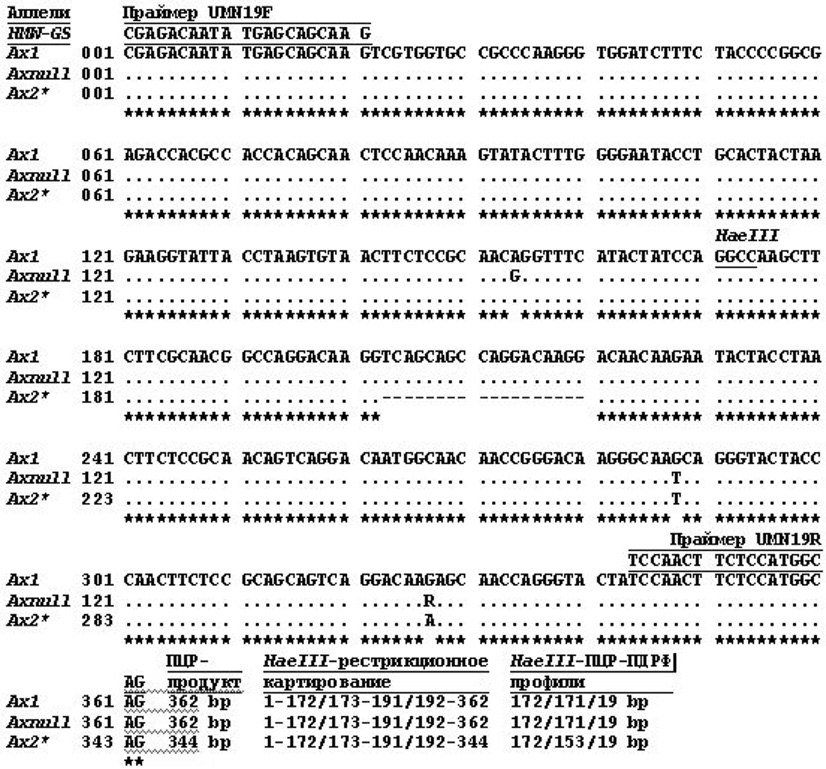
Рис. 1. Выравнивание фланкируемых с праймерами UMN19F + UMN19R нуклеотидных последовательностей Ax1-, Axnull- и Ax2*-аллелей Glu-A1-локуса HMW субъединиц глютенинов пшеницы, HaeIII-рестрикционное картирование и моделирование HaeIII-ПЦР-ПДРФ-профилей
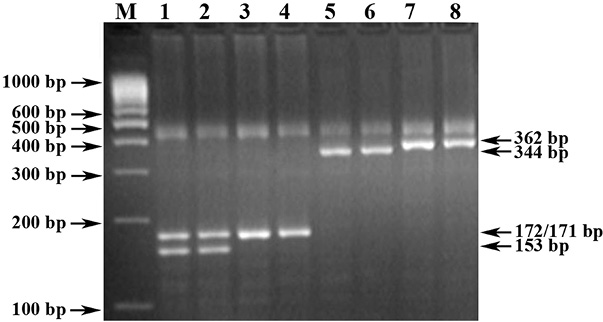
Рис. 2. Электрофореграмма технического результата предложенного способа проведения ПЦР-ПДРФ и прототипа в постановке ПЦР (праймеры UMN19F + UMN19R) для идентификации аллельных вариантов (Ax1/Axnull и Ax2*) HMW субъединиц глютенинов пшеницы
Обозначения: М) ДНК-маркеры 100 bp (СибЭнзим). 1-4) HaeIII-ПЦР-ПДРФ-анализ (предложенный способ): 1-2) Ax2*-аллель (172/153/19 bp); 3-4) Ax1/Axnull-аллели (172/171/19 bp). 5-8) ПЦР-анализ (прототип): 5-6) Ax2*-аллель (344 bp); 7-8) Ax1/Axnull-аллели (362 bp).

Рис. 3. Выравнивание фланкируемых с праймерами UMN25F + UMN25R нуклеотидных последовательностей Dx2- и Dx5-аллелей Glu-D1-локуса HMW субъединиц глютенинов пшеницы, HaeIII-рестрикционное картирование и моделирование HaeIII-ПЦР-ПДРФ-профилей
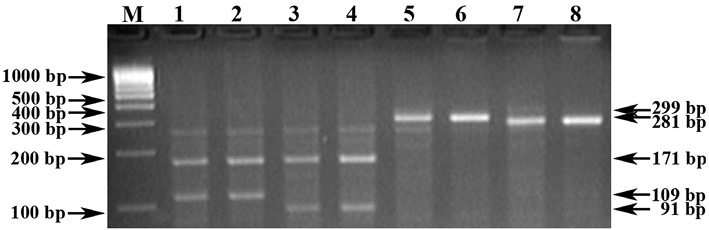
Рис. 4. Электрофореграмма технического результата предложенного способа проведения ПЦР-ПДРФ и прототипа в постановке ПЦР (праймеры UMN25F + UMN25R) для идентификации аллельных вариантов (Dx2 и Dx5) HMW субъединиц глютенинов пшеницы
Обозначения: М) ДНК-маркеры 100 bp (СибЭнзим). 1-4) HaeIII-ПЦР-ПДРФ-анализ (предложенный способ): 1-2) Dx2-аллель (171/109/19 bp); 3-4) Dx5-аллель (171/91/19 bp). 5-8) ПЦР-анализ (прототип): 5-6) Dx2-аллель (299 bp); 7-8) Dx5-аллель (281 bp).
Заключение
Учитывая общеизвестный факт положительного влияния аллельных вариантов Glu-D1d и Glu-A1b на повышение мукомольно-хлебопекарных качеств и отрицательного влияния аллельных вариантов Glu-D1a и Glu-A1a HMW-GS, приводящих к снижению хлебопекарных свойств пшеницы, можно констатировать, что из 70 исследованных образцов яровой пшеницы 32 генотипа Triticum aestivum имеют ассоциированную с высокими качествами зерна комбинацию субъединиц Ax2*/5 + 10 и могут рассматриваться как наиболее перспективные генотипы для дальнейшей селекционной работы по созданию сортов с высокими мукомольно-хлебопекарными качествами зерна.

Рис. 5. Выравнивание фланкируемых с праймерами UMN26F + UMN26R нуклеотидных последовательностей Dy10- и Dy12-аллелей Glu-D1-локуса HMW субъединиц глютенинов пшеницы, HaeIII-рестрикционное картирование и моделирование HaeIII-ПЦР-ПДРФ-профилей
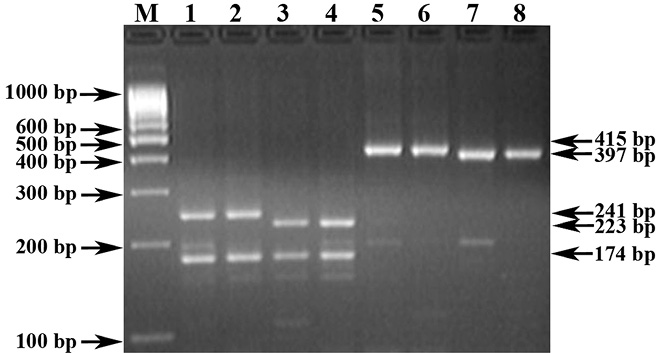
Рис. 6. Электрофореграмма технического результата предложенного способа проведения ПЦР-ПДРФ и прототипа в постановке ПЦР (праймеры UMN26F + UMN26R) для идентификации аллельных вариантов (Dy10 и Dy12) HMW субъединиц глютенинов пшеницы
Обозначения: М) ДНК-маркеры 100 bp (СибЭнзим). 1-4) HaeIII-ПЦР-ПДРФ-анализ (предложенный способ): 1-2) Dy12-аллель (241/174 bp); 3-4) Dy10-аллель (223/174 bp). 5-8) ПЦР-анализ (прототип): 5-6) Dy12-аллель (415 bp); 7-8) Dy10-аллель (397 bp).
Таблица 2
Молекулярно-генетическая оценка образцов яровой пшеницы на предмет идентификации генотипов по HMW субъединицам глютенинов
|
№ п/п |
Сорт/линия |
HMW-глютенины |
№ п/п |
Сорт/линия |
HMW-глютенины |
||||||
|
Glu-D1-локус |
Glu-A1-локус |
Glu-D1-локус |
Glu-A1-локус |
||||||||
|
5 + 10 |
2 + 12 |
Ax1 |
Ax2* |
5 + 10 |
2 + 12 |
Ax1 |
Ax2* |
||||
|
1 |
Казанская Юбилейная |
– |
+ |
– |
+ |
36 |
К-27/00-2 |
+ |
– |
– |
+ |
|
2 |
К-109/02-5 |
– |
+ |
– |
+ |
37 |
К-23/00-3 |
+ |
– |
– |
+ |
|
3 |
Экада 97 |
– |
+ |
– |
+ |
38 |
К-414/01-1 |
+ |
– |
+ |
– |
|
4 |
К-100/03-2 |
+ |
– |
– |
+ |
39 |
К-21/00 |
+ |
– |
– |
+ |
|
5 |
К-18/03-8 |
+ |
– |
– |
+ |
40 |
К-58/01-2 |
+ |
– |
– |
+ |
|
6 |
К-68/04-5 |
+ |
– |
– |
+ |
41 |
K-48/04-2 |
+ |
– |
– |
+ |
|
7 |
К-130/04-10 |
+ |
– |
– |
+ |
42 |
K-106/01-2 |
– |
+ |
– |
+ |
|
8 |
Злата |
+ |
– |
– |
+ |
43 |
K-101/04-3 |
+ |
– |
– |
+ |
|
9 |
К-88/02-19 |
+ |
– |
– |
+ |
44 |
K-112/04-2 |
+ |
– |
+ |
– |
|
10 |
К-6/01-2 |
+ |
– |
– |
+ |
45 |
K-134/04-19 |
+ |
– |
+ |
– |
|
11 |
К-5/03-6 |
– |
+ |
– |
+ |
46 |
К-51/00-3 |
+ |
– |
+ |
– |
|
12 |
К-48/03 |
+ |
– |
+ |
– |
47 |
K-133/05-5 |
+ |
– |
– |
+ |
|
13 |
К-100/03-8 |
– |
+ |
– |
+ |
48 |
K-57/05-6 |
+ |
– |
+ |
– |
|
14 |
К-21/02-5 |
– |
+ |
– |
+ |
49 |
К-117/04-4 |
+ |
– |
– |
+ |
|
15 |
К-46/04-9 |
+ |
– |
– |
+ |
50 |
К-12/04 |
– |
+ |
– |
+ |
|
16 |
К-68/04-1 |
+ |
– |
– |
+ |
51 |
К-99/05-2 |
– |
+ |
– |
+ |
|
17 |
К-23/04-1 |
+ |
– |
– |
+ |
52 |
Кк-8/06-1 |
– |
+ |
– |
+ |
|
18 |
К-49/04 |
+ |
– |
+ |
– |
53 |
Кк-71/06-3 |
+ |
– |
– |
+ |
|
19 |
К-7/04-2 |
+ |
– |
– |
+ |
54 |
Кк-8/06-6 |
– |
+ |
– |
+ |
|
20 |
Экада 113 |
– |
+ |
– |
+ |
55 |
Кк-75/06-3 |
+ |
– |
– |
+ |
|
21 |
Экада 109 |
+ |
– |
– |
+ |
56 |
Кк-11/06-11 |
+ |
– |
– |
+ |
|
22 |
К-93/05-2 |
+ |
– |
+ |
– |
57 |
Кк-11/06-10 |
+ |
– |
– |
+ |
|
23 |
К-29/02-5 |
+ |
– |
+ |
– |
58 |
Кк-69/06-4 |
– |
+ |
– |
+ |
|
24 |
К-109/02-13 |
+ |
– |
+ |
59 |
Кк-6/07-2 |
+ |
– |
– |
+ |
|
|
25 |
К-20/02-2 |
+ |
– |
– |
+ |
60 |
Кк-69/06-1 |
– |
+ |
– |
+ |
|
26 |
К-73/03-4 |
+ |
– |
– |
+ |
61 |
Кк-71/06-8 |
+ |
– |
– |
+ |
|
27 |
К-68/04-4 |
+ |
– |
– |
+ |
62 |
Кк-75/06-5 |
– |
+ |
– |
+ |
|
28 |
К-100/03-9 |
– |
+ |
– |
+ |
63 |
О-192/03-5 |
– |
+ |
– |
+ |
|
29 |
К-7/04-1 |
– |
+ |
– |
+ |
64 |
О-25/05-2 |
– |
+ |
+ |
|
|
30 |
К-65/05-2 |
+ |
– |
+ |
– |
65 |
О-206/05-2, д.515-10 |
+ |
– |
+ |
– |
|
31 |
К-11/04-8 |
– |
+ |
– |
+ |
66 |
О-186/04-1 |
+ |
– |
– |
+ |
|
32 |
Экада 66 |
+ |
– |
– |
+ |
67 |
О-464/02-2 |
– |
+ |
– |
+ |
|
33 |
Казанская Юбилейная (неполег.) |
– |
+ |
– |
+ |
68 |
О-513/00-21 |
– |
+ |
+ |
– |
|
34 |
К-65/05-1 |
+ |
– |
+ |
– |
69 |
Эр.255/00-3-1 |
– |
+ |
– |
+ |
|
35 |
Симбирцит |
+ |
– |
– |
+ |
70 |
О-28/05-2 |
– |
+ |
– |
+ |
|
Примечание: |
+ |
‒ наличие соответствующих субъединиц HMW-глютенинов |
|
– |
‒ отсутствие соответствующих субъединиц HMW-глютенинов |
Выражаем благодарность Вафину Ришаду Абдулфартовичу за оказанную финансовую поддержку.
Рецензенты:
Абрамова З.И., д.б.н., профессор кафедры биохимии Института фундаментальной медицины и биологии Казанского (Приволжского) федерального университета, г. Казань;
Багаева Т.В., д.б.н., профессор, зав. кафедрой биотехнологии Института фундаментальной медицины и биологии Казанского (Приволжского) федерального университета, г. Казань.
Работа поступила в редакцию 14.02.2013.
Библиографическая ссылка
Абдулина И.Р., Вафин Р.Р., Тюлькин С.В., Зайнуллин Л.И., Алимова Ф.К., Асхадуллин Д.Ф., Асхадуллин Д.Ф., Василова Н.З. МОЛЕКУЛЯРНАЯ ИДЕНТИФИКАЦИЯ ГЕНОТИПОВ ЯРОВОЙ ПШЕНИЦЫ ПО АЛЛЕЛЬНЫМ ВАРИАНТАМ HMW СУБЪЕДИНИЦ ГЛЮТЕНИНОВ // Фундаментальные исследования. 2013. № 4-3. С. 628-634;URL: https://fundamental-research.ru/en/article/view?id=31245 (дата обращения: 02.08.2026).