


Scientific journal
Fundamental research
ISSN 1812-7339
"Перечень" ВАК
ИФ РИНЦ = 1,798
ANATOMIC AND MORPHOMETRIC PARAMETERS OF DISTAL LIMB PARTS OF OUTBRED LABORATORY MICE AS EXPERIMENTAL MODEL FOR STUDYING OF HUMAN 2D:4D DIGIT RATIO

Рядом авторов было показано, что пальцевые индексы животных и человека взаимосвязаны с особенностями поведения, успешностью в спортивных достижениях и некоторыми физиологическими показателями [5, 7]. Предполагается, что это обусловлено сопряжёнными морфогенетическими эффектами стероидных гормонов на пренатальное развитие нервной системы и конечностей, так, что дефинитивная длина пальцев и форма кисти могут отражать особенности процессов становления структурно-функциональной организации мозга. На основании этого было предложено их использование в качестве биометрических маркеров, свидетельствующих о степени пренатальной андрогенизации структур мозга и взаимосвязанных с этим процессом показателями в зрелом периоде онтогенеза, включая гендерные предпочтения, плодовитость и риски развития отдельных заболеваний, в том числе психических [5, 7].
Разработка этой проблемы определяется необходимостью установления фундаментальных механизмов контролируемого гомеозисными генами морфогенеза конечностей и одновременно сопряжённого с этим процессом гормонально опосредованного развития мозга. С научно-практической точки зрения её актуальность диктуется поиском надёжных и простых биометрических маркёров нарушения развития и риска соответствующих патологий и поведенческих реакций. Наиболее широко известным маркёром такого рода является 2D:4D пальцевой индекс или отношение длин указательного и безымянного пальцев, известный как индекс Мэннинга [6]. Будучи психологом, Д.Т. Мэннинг в своих работах лишь высказывал гипотезу о непосредственной взаимосвязи уровня пренатального тестостерона с поведенческими особенностями взрослого человека, её правомерность экспериментально не была доказана, а полученные на плоде человека и животных данные весьма противоречивы [2, 4]. Лабораторные исследовательские модели этого феномена единичны, к настоящему времени выполнены несколько экспериментальных исследований с неоднозначными результатами [8, 9]. Одна из работ была основана на оценке эффекта введения беременным самкам лабораторных крыс этанола, снижающего уровень пренатальных андрогенов, при этом согласно полученным авторами данным, показатели пальцевого индекса остались без статистически значимых изменений [8].
Вторая работа была выполнена на развивающихся яйцах фазанов с введением различных доз тестостерона с такими же неоднозначными результатами его влияния на дефинитивную длину пальцев только левой конечности [9]. Ключевой проблемой в этом вопросе является выяснение адекватности лабораторной модели человеку и интерпретации полученных результатов. В опубликованной в 2011 году фундаментальной работе Z. Zheng и M.J. Cohn [11] был непосредственно исследован эффект пренатального введения половых стероидов на активность генов, детерминирующих рост пальцев конечностей развивающихся лабораторных мышей, митотическую активность хрящевой и костной тканей пальцевых зачатков, анатомическую длину пальцев. Ростовые эффекты стероидных гормонов в пренатальном периоде развития не могут быть адекватны аналогичным эффектам постнатального морфогенеза, по крайней мере, у человека рост отдельных костных фаланг пальцев кисти продолжается до глубокой старости и может быть обусловлен как генетическими, так и внешними факторами [1]. С анатомо-антропологической точки зрения спорным является выбор Z. Zheng и M.J. Cohn только задних конечностей для исследования. Кроме того, авторы статьи оперировали понятием длины пальца, как суммарной длины его костных сегментов. Как известно, длина костных сегментов пальца определяет его общую длину, но не идентична ей. Пальцевое отношение Мэннинга измеряется на основе общей длины пальца, включая мягкие ткани, от наиболее проксимальной сгибательной борозды у его основания, а не суммарной длины костных фаланг [5, 6]. На длину пальца влияет не только суммарная длина костных фаланг, но и его топография в составе кисти, т.е. степень выступания за пределы ладони относительно пястной кости. Топография указательного и безымянного пальцев, соотносительной длиной которых определяется в целом анатомический тип кисти и пальцевое отношение Мэннинга на 29 % определяет его значение [3] и это явление не может игнорироваться. Наконец, отсутствуют исследования, в которых бы сравнивались частоты встречаемости разных типов кисти у лабораторных животных и соответствие их аналогичным показателям человека, неизвестно как соотносятся формулы длины пальцев (пальцевые формулы) с аналогичными формулами кисти человека. Исходя из этого вопрос об адекватности использования мышей или других позвоночных в качестве лабораторной модели для изучения сопряжённого воздействия пренатальных и неонатальных стероидных гормонов на морфогенез головного мозга и конечностей, а пальцевых индексов в качестве маркеров этого процесса следует считать открытым.
Цель исследования - установление закономерностей изменчивости длины пальцев, пальцевых формул и типов конечностей лабораторных мышей как модели, адекватной человеку.
Материал и методы исследования
Настоящее исследование выполнено на 60 половозрелых аутбредных белых лабораторных мышах (28 самок и 32 самца), в возрасте 80-90 дней, средний вес самцов составил 29,2 ± 2,5 г, самок - 27,4 ± 3,1 г. Все эксперименты, уход и содержание осуществлялись в соответствии с Директивой № 63 от 22 сентября 2010 года Президиума и Парламента Европы «О защите животных, используемых для научных исследований», «Санитарными правилами по устройству, оборудованию и содержанию экспериментально-биологических клиник» от 06.04.1993 и приказом Минздрава РФ № 267 от 19.06.2003 «Об утверждении правил лабораторной практики». На проведение исследований было получено разрешение этического комитета ФГБОУ ВПО УлГУ Минобрнауки РФ. Объектом прямой оптической морфометрии послужили пальцы передних и задних конечностей животных, полученные путём ампутации их дистальных отделов в лучезапястном суставе. Кисть конечности фиксировали в 10 %-м формалине в расправленном положении в течение не менее 24 часов. Морфометрия каждого пальца проводилась с использованием стандартной окулярной измерительной вставки стереоскопического микроскопа МБС-10 при увеличениях от 5х до 16х и включала измерение его длины от середины проксимальной сгибательной кожной борозды до дистальной точки (dactilion) со стороны ладонной поверхности [4]. Палец измеряли в выпрямленном состоянии, придерживая его за ноготь на предметном столике препаровальной иглой (рисунок, а). Для определения значения деления шкалы морфометрической окулярной вставки при каждом увеличении использовали объект-микрометр «ОМ-П» фирмы «ЛОМО» ГОСТ 7513-56 с делением шкалы 0,01 мм. Тип кисти определяли по дистальному профилю соотношения кончиков II и IV пальцев [4]. Статистическую обработку данных проводили с использованием лицензионной компьютерной программы «Statistica 6.0» StatSoftInc. (США) с использованием параметрической статистики по правилам, рекомендованным международным комитетом редакторов биомедицинских журналов (ICMJE). Для попарных сравнений средних величин использовали t-критерий. При проведении апостериорных множественных сравнений ANOVA использовали критерий Бонферрони.
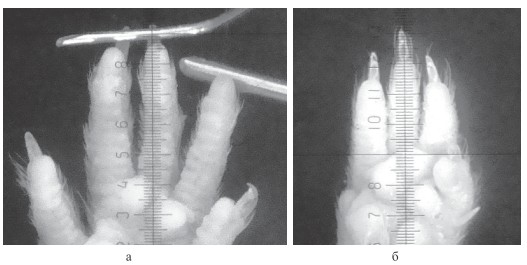
Микрофото дистальных отделов конечностей белой лабораторной мыши: а - правая задняя конечность в расправленном состоянии с подошвенной стороны; б - левая передняя конечность с ладонной стороны. Ув. х16
Результаты исследования и их обсуждение
Определение границ изменчивости изучаемого параметра для животных одного пола производилось в следующих направлениях: унилатеральном (для каждой конечности отдельно), билатеральном (попарно, для передних, задних), краниокаудальном (попарно, ипсилатерально).
Кроме того, были произведены попарные сопоставления для каждой конечности у животных разного пола. У исследованных нами животных на передней конечности первые пальцы были редуцированы и в исследование не включались, но учитывались в пальцевой формуле (рисунок, б). Согласно полученным результатам, по среднему значению длины на передней правой конечности у самцов преобладает III палец (2,79 ± 0,22 мм, таблица).
Однофакторный дисперсионный анализ позволил получить следующую пальцевую формулу длины для этой конечности: III > IV > II > V > I (p < 0,05).На левой передней конечности самцов разница между длиной II и IV пальцев оказалась статистически не значимой, в результате пальцевая формула для этой конечности имела вид: III > II = IV > V > I, при значимых различиях показателей III и V пальцев между собой и значениями IV и II-го. На обеих задних конечностях самцов в длине II, III, IV пальцев нами не выявлено статистически значимых различий, пальцевая формула имела следующий вид: II = III = IV > V > I. Билатеральное сравнение длины пальцев передних конечностей не обнаружило достоверных различий. При аналогичном сравнении длин пальцев задних конечностей статистически значимой оказалась разница в длине V-х пальцев. На правой задней конечности длина V пальца преобладала над длиной слева - 3,26 ± 0,27 и 3,20 ± 0,26 мм, соответственно (p < 0,05; таблица). Наконец, при краниокаудальном сравнении длин пальцев у самцов статистически значимо более высокие показатели длины пальцев выявлены на задней конечности, как с левой, так и с правой стороны тела (см. таблицу). Все перечисленные выше особенности изменчивости длины пальцев при унилатеральном и билатеральном сравнениях, присущие для животных мужского пола, равным образом характерны и для животных женского пола (см. таблицу). При краниокаудальном сравнении выявились небольшие половые отличия, заключающиеся в том, что у самок длина одноимённых II и V пальцев передних и задних конечностей на правой стороне тела статистически значимо не различались, в то время как у самцов на задних конечностях она преобладала на всех пальцах без исключения. На левой стороне тела, так же как у самцов, длины пальцев задней конечности преобладали над показателями передней конечности (см. таблицу). Нами установлено, что дистальные профили всех конечностей исследованных популяции лабораторных мышей по соотносительной длине выступания II и IV пальцев, по которым у человека определяется тип кисти, соответствуют так называемому ульнарному (у человека - преимущественно мужскому) типу кисти, в котором кончик IV пальца выступает дальше кончика II (см. рисунок, а). Из 60 наблюдений лишь в двух случаях наблюдался промежуточный или неопределённый тип дистального профиля конечности, выявленный нами как артефакт травмированного IV пальца (см. рисунок, б).
Длина пальцев, мм, конечностей самцов (n = 32, ♂) и самок (n = 28, ♀) белых беспородных лабораторных мышей (M ± σ)
|
Конечность |
Пол |
Номера пальцев |
||||
|
I |
II |
III |
IV |
V |
||
|
Левая передняя |
♂ |
- |
2,47 ± 0,20 |
2,77 ± 0,19 |
2,58 ± 0,29 |
2,01 ± 0,20 |
|
♀ |
- |
2,47 ± 0,16 |
2,75 ± 0,19 |
2,64 ± 0,18 |
1,98 ± 0,18 |
|
|
Правая передняя |
♂ |
- |
2,46 ± 0,22 |
2,79 ± 0,22 |
2,61 ± 0,23 |
2,02 ± 0,22 |
|
♀ |
- |
2,46 ± 0,19 |
2,80 ± 0,16 |
2,59 ± 0,21 |
2,05 ± 0,19 |
|
|
Левая задняя |
♂ |
2,22 ± 0,28 |
3,62 ± 0,28 |
3,71 ± 0,28 |
3,68 ± 0,28 |
3,20 ± 0,26 |
|
♀ |
2,16 ± 0,16 |
3,69 ± 0,24 |
3,70 ± 0,25 |
3,63 ± 0,27 |
3,20 ± 0,22 |
|
|
Правая задняя |
♂ |
2,16 ± 0,19 |
3,62 ± 0,29 |
3,76 ± 0,34 |
3,70 ± 0,31 |
3,26 ± 0,27 |
|
♀ |
2,12 ± 0,17 |
3,67 ± 0,22 |
3,68 ± 0,23 |
3,59 ± 0,25 |
3,22 ± 0,22 |
|
Пальцевая формула длины пальцев кисти человека, независимо от пола, выглядит следующим образом: III > II = IV > I > V [3]. С учётом редукции у мышей первых пальцев на передних конечностях наиболее соответствующей аналогичной пальцевой формуле человека, следовало бы признать пальцевую формулу передней левой конечности самцов: III > II = IV > V > I. Исходя из того, что 2D:4D пальцевой индекс был Мэннингом впервые использован для лиц мужского пола, то, на наш взгляд, следует считать наиболее адекватной для оценки экспериментальных воздействий признать не левую, а правую переднюю конечность. Длина IV пальца этой конечности по своим средним значениям превышает длину II пальца. Кроме того, по морфологическому типу и пальцевой формуле, она фактически является конечностью ульнарного, то есть преимущественно мужского типа.
При рассмотрении вопроса об адекватности закономерностей изменчивости пальцевых формул лабораторных мышей аналогичным закономерностям у человека следует принимать во внимание, как соотношение абсолютных значений морфометрических параметров, так и значение ряда функциональных факторов [3]. Выбор некоторыми исследователями для оценки эффектов гормонов на рост пальцев в период пренатального онтогенеза задних конечностей [11], имеющих у взрослых особей формулу II = III = IV > V > I, может быть оправдан только размерными анатомическими, но не функциональными или морфогенетическими соображениями. Передние конечности у большинства высокоорганизованных позвоночных получают в пренатальном онтогенезе преимущественное развитие и обладают более высокой степенью дифференцировки и специализации, связанной, в том числе, с развитием мозга. Более адекватные и аргументированно интерпретируемые экспериментальные результаты по пренатальной андрогенизации были получены раньше, чем в работе Z. Zheng и M.J. Cohn, на передних конечностях крыс линии Вистар [10]. Из-за отсутствия сравнительно-анатомических данных по пальцевым формулам с учётом билатеральной организации и функциональной специализации (не исключая влияния генетических факторов) передних конечностей крыс и человека, авторы не смогли объяснить феномен преимущественных изменений на левой, но не правой конечности. У человека процесс формирования кисти и рост пальцев продолжается на протяжении всего онтогенеза [1], использование для пальцевого индекса соотношения длин пальцев двух основных (II и IV) морфогенетических лучей кисти, а не стопы, в этом отношении сомнений не вызывает. Задние конечности животного по сравнению с передними в постнатальном онтогенезе не испытывают влияний латерализации, функциональных факторов, связанных с манипуляциями приёма пищи, исследовательскими движениями, поведенческими реакциями. В этом смысле они лишены возмущающего воздействия и, безусловно, могут лучше отражать влияние гормонов в раннем онтогенезе, структурно более устойчивое и проявляющееся в дефинитивной соотносительной длине пальцев. Исследование Z. Zheng и M.J. Cohn [11] лишь доказывает дифференцированное влияние стероидов на рост костных структур отдельных пальцев задней конечности в целом и не может служить полным аналогом гормонально зависимого, в значительной степени функционально опосредованного и изменяющегося с возрастом роста пальцев кисти человека.
Заключение
Таким образом, полученные формулы длины пальцев конечностей мышей в целом имеют однотипное сходство с аналогичной формулой кисти человека, в особенности правая передняя конечность у самцов. Несмотря на некоторую разницу в анатомическом строении дистальных отделов конечностей человека и лабораторной мыши, отсутствие абсолютной идентичности формул длины пальцев, полученные данные доказывают возможность и адекватность использования лабораторных аутбредных мышей для изучения влияния пренатального введения стероидных гормонов на морфогенез пальцев кисти.
Рецензенты:
Музурова Л.В., д.м.н., профессор кафедры анатомии человека ГОУ ВПО «Саратовский ГМУ имени В.И. Разумовского», г. Саратов;
Чарышкин А.Л., д.м.н., профессор, заведующий кафедрой факультетской хирургии ФГБОУ ВПО «Ульяновский государственный университет», г. Ульяновск.
Работа поступила в редакцию 12.12.2012.
Библиографическая ссылка
Фомина А.В., Хайруллин Р.М., Зеркалова Ю.Ф. АНАТОМО-МОРФОМЕТРИЧЕСКАЯ ХАРАКТЕРИСТИКА ДИСТАЛЬНЫХ ОТДЕЛОВ КОНЕЧНОСТЕЙ ЛАБОРАТОРНЫХ МЫШЕЙ КАК ЭКСПЕРИМЕНТАЛЬНОЙ МОДЕЛИ ДЛЯ ИССЛЕДОВАНИЯ 2D:4D ПАЛЬЦЕВОГО ИНДЕКСА ЧЕЛОВЕКА // Фундаментальные исследования. 2012. № 12-2. С. 381-385;URL: https://fundamental-research.ru/en/article/view?id=30905 (дата обращения: 01.07.2026).