


Scientific journal
Fundamental research
ISSN 1812-7339
"Перечень" ВАК
ИФ РИНЦ = 1,798
MOLECULAR IDENTIFICATION OF SPRING WHEAT GENOTYPES BY ALLELIC VARIANTS OF THE WAXY-GENES

Крахмал, являющийся основным компонентом зерновки пшеницы, представлен полисахаридами двух типов – разветвленного амилопектина и линейной амилозы, соотношение которых предопределяет технологические качества крахмала с тенденцией к улучшению мукомольно-хлебопекарных и технологических свойств зерна при снижении концентрации амилозы с 20 до 0 % [1, 2, 3, 4, 5, 6].
Амилоза крахмала синтезируется за счет активности гранул-связанной синтазы крахмала (GBSSI), кодируемой 3 генами, получивших название Waxy, чьи b-аллельные варианты локусов, являющиеся нефункциональными нуль-аллелями, влияют на образование крахмала с пониженным содержанием амилозы (при наличии одного или двух нуль-аллелей), и состоящего только из амилопектина (при наличии всех трех нуль-аллелей) [1, 2, 3, 4, 5].
Наиболее перспективными подходами к оценке аллельного полиморфизма Waxy-генов являются способы идентификации на основе молекулярно-генетических методов исследования – ключевого инструментария в маркер-вспомогательной селекции [1, 2, 3, 4, 5].
Целью настоящей работы являлась молекулярная идентификация перспективных генотипов яровой пшеницы селекции ТатНИИСХ по аллельным вариантам Waxy-генов для создания сортов с высокими мукомольно-хлебопекарными и технологическими свойствами зерна.
Материалы и методы исследования
Молекулярно-генетическая оценка 70 образцов яровой пшеницы преимущественно селекции ТатНИИСХ на предмет идентификации генотипов по аллельным вариантам Waxy-генов проведена методами ПЦР- и ПЦР-ПДРФ-анализа на основе общепринятых [4, 5] и разработанных нами способов генотипирования с дополнительным обоснованием достоверности полученных результатов исследования методом прямого секвенирования амплифицированных фрагментов ДНК.
Экстракция геномной ДНК из зерновок растений яровой пшеницы молочно-восковой спелости генерации 2012 г. осуществлена коммерческим набором «ДНК-сорб С» («ЦНИИ эпидемиологии», Россия). Амплификация геномной ДНК проведена на термоциклерах «Терцик» («ДНК-технология», Россия) и «MyCycler» с градиентом («Bio-Rad», США) с использованием олигонуклеотидных праймеров, перечень которых представлен в таблице.
Условия проведения ПЦР- и ПЦР-ПДРФ-анализа для идентификации аллельных вариантов Waxy-генов пшеницы
|
Праймеры |
Последовательности праймеров (5/-3/) |
Локус |
Режим амплификации |
ПДРФ-анализ |
|
4F |
AAGAGCAACTACCAGT |
Wx-A1 Wx-B1 Wx-D1 |
×1: 94 °С – 4 мин ×40: 94 °С – 30 с, 58 °С – 30 с, 72 °С – 30 с ×1: 72 °С – 7 мин |
|
|
4R |
TCGTACCCGTCGATGAAGTCGA |
|||
|
4F-c |
CCCCCAAGAGCAACTACCAGT |
Wx-A1 Wx-B1 Wx-D1 |
×1: 94 °С – 4 мин ×40: 94 °С – 30 с, 64 °С – 30 с, 72 °С – 30 с ×1: 72 °С – 7 мин |
AcsI 50 °C – 3 ч |
|
4R |
TCGTACCCGTCGATGAAGTCGA |
|||
|
Wx-A1L: |
CCCCAAAGCAAAGCAGGAAAC |
Wx-A1 |
×1: 94 °С – 4 мин ×40: 94 °С – 45 с, 55 °С – 30 с, 72 °С – 1 мин ×1: 72 °С – 7 мин |
HindIII 37 °C – 3 ч |
|
Wx-A1R |
CGGCGTCGGGTCCATAGATC |
Детекция результатов ПЦР- и ПЦР-ПДРФ-анализа выполнена методом горизонтального электрофореза в 2–3 % агарозном геле в буфере ТBE (рН 8,0), содержащем этидий бромид с последующей визуализацией результатов в ультрафиолетовом трансиллюминаторе (λ = 310 нм) гельдокументирующей системы Gel Doc (Bio-Rad, США).
Размеры фрагментов ДНК оценены по подвижности в сравнении со стандартными ДНК маркерами. В работе использованы реактивы для молекулярно-биологических исследований производства ООО «СибЭнзим» (Россия).
Секвенирование продуктов амплификации проведено на генетическом анализаторе «ABI PRISM 3500» в НПО «Синтол» (Россия). Выравнивание секвенируемых последовательностей ДНК осуществлено с использованием программы BLAST NCBI.
Результаты исследования и их обсуждение
По результатам молекулярной идентификации генотипов по аллельным вариантам Waxy-генов установлено, что из 70 происследованных образцов яровой пшеницы 65 растений (92,8 %) имели комбинацию активных аллелей Wx-A1a/B1a/D1a (1-ый дикий тип), 3 образца: Кк-11/06-11, Кк-11/06-10 и Кк-69/06-1 (4,4 %) – Wx-A1g/B1a/D1a (неклассифицированный тип), и лишь 2 линии: Кк-8/06-6 и О-192/03-5 (2,8 %) по классификации типов пшеницы с различным содержанием Wx-генов относились к 3-му типу (Wx-A1a/B1b/D1a) (табл. 2).
Для эффективной аллельной дискриминации Wx-A1g и Wx-B1e от нуль-аллелей Wx-A1b и Wx-B1b соответственно нами были разработаны оригинальные способы генотипирования, повышающие точность ДНК-анализа, с дополнительным обоснованием достоверности тестов секвенированием ПЦР-продуктов и депонированием расшифрованных нуклеотидных последовательностей в GenBank NCBI (GenBank A/N: JX649155-JX649158).
Оригинальность предложенного нами способа генотипирования в части аллельной дискриминации Wx-A1g от нуль-аллеля Wx-A1b заключается в реконструкции прямого праймера 4F [4, 5] путем наращивания его 5/-концевого участка пятизвенным oligo (dC)5 блоком для выравнивания температур плавлений реконструированного (4F-c) и обратного (4R) праймеров и подбора оптимальной температуры отжига (Ta = 64 °C). Сам же принцип дискриминации данных аллелей основан на учете как наличия амплификации фрагмента локуса Wx-A1g размером 257 bp разной степени интенсивности сигнала в постановке ПЦР с праймерами 4F + 4R [4, 5] (рис. 1, а, треки 5–7), так и отсутствия ПЦР-продукта размером 262 bp в предложенном нами способе генотипирования с олигонуклеотидами 4F-c + 4R (рис. 1, в, треки 5–7).
Помимо обоснования достоверности ДНК-тестов секвенированием ПЦР-продуктов дополнительно была проведена процедура ПЦР-ПДРФ-анализа по Wx-A1-локусу с праймерами Wx-A1L + Wx-A1R и эндонуклеазным расщеплением рестриктазой HindIII [5], также подтвердившая правильность интерпретации выявленных аллелей.
Оригинальность же разработанного нами способа генотипирования в части дискриминации нуль-аллеля Wx-B1b от активного Wx-B1e-аллеля заключается в подборе условий ПЦР-ПДРФ-анализа с праймерами 4F-c + 4R и эндонуклеазным расщеплением рестриктазой AcsI, генерирующих генотипические профили, в частности, с характерным для Wx-B1b-аллеля отсутствием ПДРФ-фрагмента длиной 84 bp (рис. 2, треки 6–7), или наличием неразрезанного ПЦР-продукта локуса Wx-B1e-аллеля размером 266 bp в виду отсутствия у него соответствующего сайта рестрикции (R↑AATTY).
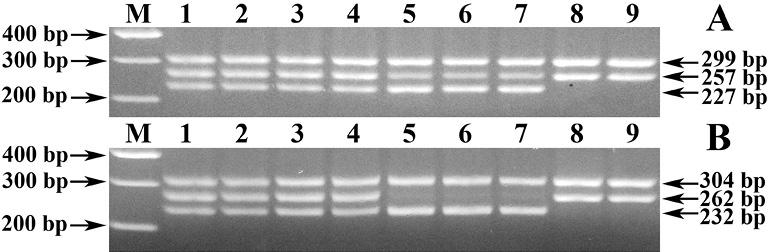
Рис. 1. Электрофореграмма результата ПЦР-идентификации генотипов яровой пшеницы селекции ТатНИИСХ по аллельным вариантам Waxy-генов. Обозначения: результаты ПЦР-идентификации с праймерами 4F + 4R (А) и 4F-c + 4R (B). М – ДНК-маркеры 100–1500 bp (СибЭнзим). 1–9 – генотипы с комбинацией Wx-аллелей: 1–4 – Wx-A1a/B1a/D1a; 5–7 – Wx-A1g/B1a/D1a. 8–9 – Wx-A1a/B1b/D1a
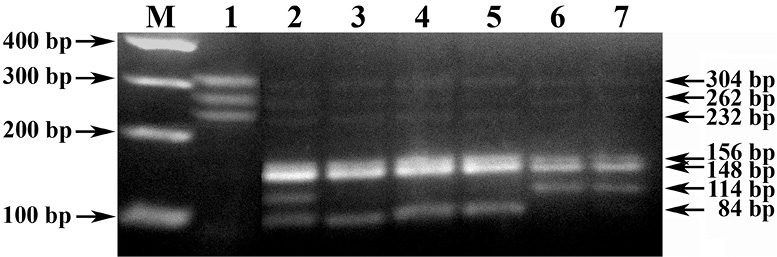
Рис. 2. Электрофореграмма результата ПЦР-ПДРФ-идентификации генотипов яровой пшеницы селекции ТатНИИСХ по аллельным вариантам Waxy-генов. Обозначения: М – ДНК-маркеры 50–100 bp (СибЭнзим). 1 – ПЦР-профиль генотипа с комбинацией аллелей Wx-A1a/B1a/D1a (304/262/232 bp). 2–7 – AcsI-ПДРФ-профиль генотипов с комбинацией Wx-аллелей: 2 – Wx-A1a/B1a/D1a (156/148/114/84 bp); 3–5 – Wx-A1g/B1a/D1a (156/148/84 bp); 6–7 – Wx-A1a/B1b/D1a (156/148/114 bp)
Таким образом, из общего числа проанализированных образцов растений (табл. 2) наиболее перспективными генотипами, рассматриваемыми в качестве исходного материала для дальнейшей селекционной работы по созданию сортов яровой пшеницы с крахмалом амилопектинового типа (с низким содержанием амилозы), являются две линии Кк-8/06-6 и О-192/03-5, несущие в своем геноме нулевой Wx-B1b-аллель, с последующим введением Wx-A1b- и (или) Wx-D1b-аллеля путем скрещивания с донорами нуль-аллелей по локусам Waxy-генов.
Заключение
Представленная в настоящей работе информация является фактически первым опубликованным в виде научной статьи упоминанием о достоверном выявлении аллельных вариантов Wx-A1g и Wx-B1b Waxy-генов у генотипов яровой пшеницы отечественной селекции. Разработанные нами способы генотипирования по аллельным вариантам Waxy-генов повышают достоверность идентификации в части, касающейся дискриминации активных аллелей Wx-A1g и Wx-B1e от нуль-аллелей Wx-A1b и Wx-B1b.
Таблица 2
Молекулярно-генетическая оценка образцов яровой пшеницы на предмет идентификации генотипов по аллельным вариантам Waxy-генов
|
№ п/п |
Сорт/линия |
Waxy-гены |
№ п/п |
Сорт/линия |
Waxy-гены |
||||||||||||||
|
A1 |
B1 |
D1 |
A1 |
B1 |
D1 |
||||||||||||||
|
a |
b |
g |
a |
b |
e |
a |
b |
a |
b |
g |
a |
b |
e |
a |
b |
||||
|
1 |
Казанская Юбилейная |
+ |
– |
– |
+ |
– |
– |
+ |
– |
36 |
К-27/00-2 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
|
2 |
К-109/02-5 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
37 |
К-23/00-3 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
|
3 |
Экада 97 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
38 |
К-414/01-1 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
|
4 |
К-100/03-2 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
39 |
К-21/00 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
|
5 |
К-18/03-8 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
40 |
К-58/01-2 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
|
6 |
К-68/04-5 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
41 |
K-48/04-2 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
|
7 |
К-130/04-10 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
42 |
K-106/01-2 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
|
8 |
Злата |
+ |
– |
– |
+ |
– |
– |
+ |
– |
43 |
K-101/04-3 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
|
9 |
К-88/02-19 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
44 |
K-112/04-2 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
|
10 |
К-6/01-2 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
45 |
K-134/04-19 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
|
11 |
К-5/03-6 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
46 |
К-51/00-3 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
|
12 |
К-48/03 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
47 |
K-133/05-5 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
|
13 |
К-100/03-8 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
48 |
K-57/05-6 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
|
14 |
К-21/02-5 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
49 |
К-117/04-4 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
|
15 |
К-46/04-9 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
50 |
К-12/04 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
|
16 |
К-68/04-1 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
51 |
К-99/05-2 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
|
17 |
К-23/04-1 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
52 |
Кк-8/06-1 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
|
18 |
К-49/04 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
53 |
Кк-71/06-3 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
|
19 |
К-7/04-2 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
54 |
Кк-8/06-6 |
+ |
– |
– |
– |
+ |
– |
+ |
– |
|
20 |
Экада 113 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
55 |
Кк-75/06-3 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
|
21 |
Экада 109 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
56 |
Кк-11/06-11 |
– |
– |
+ |
+ |
– |
– |
+ |
– |
|
22 |
К-93/05-2 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
57 |
Кк-11/06-10 |
– |
– |
+ |
+ |
– |
– |
+ |
– |
|
23 |
К-29/02-5 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
58 |
Кк-69/06-4 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
|
24 |
К-109/02-13 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
59 |
Кк-6/07-2 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
|
25 |
К-20/02-2 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
60 |
Кк-69/06-1 |
– |
– |
+ |
+ |
– |
– |
+ |
– |
|
26 |
К-73/03-4 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
61 |
Кк-71/06-8 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
|
27 |
К-68/04-4 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
62 |
Кк-75/06-5 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
|
28 |
К-100/03-9 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
63 |
О-192/03-5 |
+ |
– |
– |
– |
+ |
– |
+ |
– |
|
29 |
К-7/04-1 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
64 |
О-25/05-2 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
|
30 |
К-65/05-2 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
65 |
О-206/05-2, д.515-10 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
|
31 |
К-11/04-8 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
66 |
О-186/04-1 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
|
32 |
Экада 66 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
67 |
О-464/02-2 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
|
33 |
Казанская юбилейная (неполег.) |
+ |
– |
– |
+ |
– |
– |
+ |
– |
68 |
О-513/00-21 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
|
34 |
К-65/05-1 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
69 |
Эр.255/00-3-1 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
|
35 |
Симбирцит |
+ |
– |
– |
+ |
– |
– |
+ |
– |
70 |
О-28/05-2 |
+ |
– |
– |
+ |
– |
– |
+ |
– |
Примечания:
+ – наличие соответствующих аллелей Waxy-генов;
– – отсутствие соответствующих аллелей Waxy-генов.
Рецензенты:
Абрамова З.И., д.б.н., профессор кафедры биохимии Института фундаментальной медицины и биологии Казанского (Приволжского) федерального университета, г. Казань;
Багаева Т.В., д.б.н., профессор, зав. кафедрой биотехнологии Института фундаментальной медицины и биологии Казанского (Приволжского) федерального университета, г. Казань.
Работа поступила в редакцию 26.11.2012.
Библиографическая ссылка
Абдулина И.Р., Вафин Р.Р., Ржанова И.В., Гараева А.Л., Асхадуллин Д.Ф., Асхадуллин Д.Ф., Василова Н.З., Зайнуллин Л.И., Алимова Ф.К. МОЛЕКУЛЯРНАЯ ИДЕНТИФИКАЦИЯ ГЕНОТИПОВ ЯРОВОЙ ПШЕНИЦЫ ПО АЛЛЕЛЬНЫМ ВАРИАНТАМ WAXY-ГЕНОВ // Фундаментальные исследования. 2013. № 1-1. С. 13-17;URL: https://fundamental-research.ru/en/article/view?id=30861 (дата обращения: 01.07.2026).