


Scientific journal
Fundamental research
ISSN 1812-7339
"Перечень" ВАК
ИФ РИНЦ = 1,798
Influence abiotic factors on transfor Vibrio cholerae to nonculturable form

Обнаружение некультивируемого состояния (НС) у бактерий обусловило необходимость нового подхода к анализу экологических и молекулярно-генетических механизмов существования возбудителей инфекций в природе.
Знание этих механизмов необходимо для оценки факторов, обеспечивающих микроорганизмам существование в качестве естественных компонентов природных биоценозов, что, в свою очередь, будет способствовать пониманию природы эндемичности и резервации патогенных бактерий в межэпизоотические (межэпидемические) периоды.
Считается, что переход бактерий в НС является ответом на ухудшение условий обитания. К основным факторам индуцирующим переход в НС бактерий исследователи относят понижение температуры в среде обитания и условия голодания (4, 7, 9).
В связи с важностью изучения экологических аспектов перехода холерных вибрионов в некультивируемое состояние особого внимания заслуживают работы, посвященные роли биотических и абиотических факторов водоёмов в этом процессе. Имеются сведения о переходе вибрионов, в том числе холерных, в некультивируемую форму (НФ) в условиях пониженной (+4 ºС) и повышенной (37 ºС) температуры (3,8), получены НФ холерных вибрионов в голодной среде, морской, речной воде и некоторых других модельных системах (5,6).
Цель настоящей работы - изучение раздельного и комплексного влияния некоторых абиотических экологических факторов: температуры, освещенности и различной солености среды на переход в некультивируемое состояние токсигенных и нетоксигенных холерных вибрионов.
Материалы и методы. В работе было использовано 9 типичных по родовым и видовым свойствам ctx+ и ctx- штаммов V. cholerae О1 и О139 серогруппы. Для получения НФ конструировали микрокосмы на основе автоклавированной речной, морской (Азовского и Черного моря) воды. В качестве контроля использовали дистиллированную воду и 0,85% NaCl. Для облегчения визуального контроля морфологии и подсчета клеток под микроскопом концентрацию холерных вибрионов доводили средой культивирования до 109 кл/мл. Пробы содержали при температуре 4-6 и 8-10 0 С без освещения и в условиях постоянной искусственной освещенности в 300 люкс.
Высевы на агаровые среды производили ежедневно в течение 30 дней, затем раз в неделю и в месяц. Некультивируемыми считали вибрионы в пробе, в высевах из которой на агаровых средах не обнаружено выросших колоний, но витальными методами окрашивания выявлены живые бактериальные клетки или ПЦР методом - ДНК ctx гена при культивировании токсигенных штаммов.
Полученные материалы проанализированы с учетом влияния на скорость перехода холерных вибрионов в НС раздельного и сочетанного действия двух и более факторов.
Результаты и обсуждение
Использование в экспериментах разного температурного режима содержания микрокосм показало, что наиболее быстрый переход холерных вибрионов в НС происходил при температуре 4-6 0С, как при постоянном искусственном освещении, так и без него.
В микрокосмах с речной водой при температуре 4-6 0С без искусственного освещения штаммы V. cholerae О1 переходили в НС через 12 - 15 суток, V. cholerae О139 - в период от 30 до 50-60 суток. В микрокосмах с водой Азовского моря НФ холерных вибрионов О1 серогруппы получены в течение 60 суток, а О 139 серогруппы - 75-90. В воде из Черного моря в тех - же условиях штаммы V. cholerae О1 переходили в НС в более поздние сроки (90 - 150 суток), а холерные вибрионы О139 серогруппы в течении 180 суток сохраняли жизнеспособность (срок наблюдения). В 0,85% растворе NaCl время перехода в НС колебалось у V. cholerae О1 от 30 до 40, а у холерных вибрионов О139 серогруппы - от 40 до 75 суток. В дистиллированной воде штаммы V. cholerae О1 переходили в НС через 30 - 40 суток, а холерные вибрионы О139 серогруппы в среднем через 45 суток, однако имели место и более поздние (до 120 суток) сроки.
В условиях постоянного искусственного освещения в микрокосмах с речной водой при температуре 4-6 0С время перехода в НС, в отличие от аналогичных микрокосм без освещённости, было значительно более продолжительным и составило для V. cholerae О1 90 суток, а для холерных вибрионов О139 серогруппы - 120-180 суток. В микрокосмах с водой Азовского моря V. cholerae О1 переходили в НС через 120 суток, а холерные вибрионы О139 серогруппы - в течение 100-150. В воде Чёрного моря V. cholerae О1 в течение 190-210 суток сохраняли жизнеспособность, а холерные вибрионы О139 серогруппы переходили в НС через 120 - 270 суток. Аналогичная тенденция отмечена и для проб с 0,85% раствором NaCl, где при постоянном искусственном освещении V. cholerae О1 переходили в НФ через 120-130 суток, а холерные вибрионы О139 серогруппы - 120 - 150 суток. В дистиллированной воде время перехода в НС V. cholerae О1, так же, как и без освещения, колебалось в пределах 30 - 45 суток, а холерных вибрионов О139 серогруппы - 60 суток.
Повышение температуры культивирования до 8-10 0С также выявило увеличение сроков перехода в НС холерных вибрионов во всех микрокосмах кроме дистиллированной воды.
Культивирование холерных вибрионов при 8-10 0С без искусственного освещения в микрокосмах с речной водой показало, что штаммы V. cholerae О1 переходили в НС через 150 - 180 суток, а холерные вибрионы О139 серогруппы - 240 - 300 суток. В микрокосмах с водой из Черного моря V. cholerae О1 и холерные вибрионы О139 серогруппы образовывали в НФ через 210 - 230 суток, в 0,85% растворе NaCl V. cholerae О1 - через 75 - 80 суток, а холерные вибрионы О139 серогруппы -114 - 120 суток. В дистиллированной воде V. cholerae О1 и V. cholerae О139 переходили в НС через 30 - 40 суток.
Освещение микрокосм и при 8-10 0С значительно увеличило время перехода холерных вибрионов в НС во всех средах за исключением дистиллированной воды. Так, при культивировании микрокосм с речной водой в условиях освещения при температуре 8-10 0С V. cholerae О1 переходили в НС через 240 - 260 суток, а холерные вибрионы О139 серогруппы - 290 - 360 суток. В микрокосмах с водой из Черного моря V. cholerae О1 переходили в НС через 275-280, а О139 серогруппы - через 280 - 340 суток. В 0,85% растворе NaCl время перехода V. cholerae О1 занимало 240 - 260 суток, а для холерных вибрионов О139 серогруппы составило 250 - 270 суток. Скорость перехода холерных вибрионов в НС при 8-10 0С в дистиллированной воде практически не изменилась: V. cholerae О1 переходили в НС через 28 - 45 суток, а холерные вибрионы О139 серогруппы - через 30 суток.
Таким образом, несмотря на наиболее короткие сроки перехода при температуре 4-6 0С холерных вибрионов в НС выявлено влияние на их продолжительность освещенности и состава среды. Например, в микрокосмах речной воды, соленость которой гораздо ниже, чем из Черного моря, при постоянном искусственном освещении время перехода V. cholerae О1 в НС оказывается в 6,8, а V. cholerae О139 - в 3,7 раза больше, чем без неё. В микрокосмах с водой Азовского моря освещённость в среднем в 1,7 раза увеличивала эти сроки, а в воде из Черного моря - в 1,2 раза. При культивировании холерных вибрионов в 0,85% растворе NaCl, сроки перехода в НС в условиях освещенности увеличивались в 2,3 раза. В дистиллированной воде влияния освещённости на переход холерных вибрионов в НС не выявлено, ни при 4-6, ни при 8-10 0С.
В условиях микрокосм с различной соленостью среды, температурой содержания и освещенностью не выявлено зависимости перехода в НС холерных вибрионов от их токсигенности. Обнаружено лишь различие в скорости образования НФ холерными вибрионами разных серогрупп. Различия в скорости перехода в НС V.cholerae О1 и О139 особенно выражены в микрокосмах с морской и речной водой. Так например, V. cholerae О1 в микрокосмах с речной водой без освещения в 3,1 раза быстрее переходили в НС, чем V. cholerae О139, в условиях освещенности сроки перехода в НС у V. cholerae О139 увеличились в 1,2 - 1,7 раза (Рисунок).
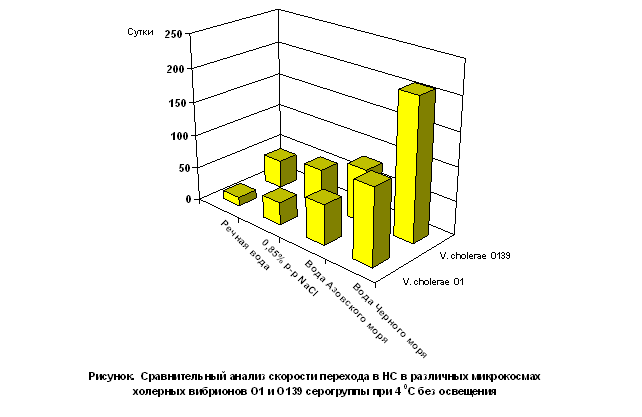
В микрокосмах с водой из Чёрного моря продолжительность сохранения в культивируемом состояний V. cholerae О139 серогруппы была в 1,7 раза больше, чем у V. cholerae О1. При искусственном освещении у V. cholerae О1 переход в НФ в воде из Черного моря не зарегистрирован, а сроки образования НФ у V. cholerae О139 были более продолжительными чем без освещения.
Получение в экспериментальных экосистемах НФ холерных вибрионов под действием нескольких факторов, характеризующих природные экосистемы, выявило достоверно значимое влияние на скорость процесса как каждого из изученных факторов, так и совместного их действия в различных сочетаниях.
Из четырех изученных микрокосм с различным солевым составом среды в условиях без искусственной освещенности при 4-6 ºС в трех (кроме морской воды, характеризующейся более высоким, чем в остальных микрокосмах, содержанием солей) холерные вибрионы переходили в НС в одинаковые сроки. Освещенность этих же микрокосм увеличивала сроки перехода в НС холерных вибрионов во всех четырех микрокосмах: в 0,85% растворе NaCl в - 2,3 раза, в речной воде в 4,1 раза и в воде Азовского моря - в 1,7 раза. В морской воде, при искусственном освещении НФ V. cholerae О1 не были получены, в отличие от аналогичных микрокосм без освещения, где холерные вибрионы перешли в НС в течение 190-210 суток.
При температуре 8-10 0С в тех же микрокосмах без искусственного освещения сроки перехода в НС удлинялись: в речной воде - в 7,4 раза, в 0,85% растворе NaCl - в 1,8 раза и в морской воде в - 1,6 раза. Искусственное освещение в условиях этой температуры еще более увеличило сроки перехода в НС холерных вибрионов: в 0,85% растворе NaCl в 2,8 раза, в речной воде в 1,3 раза и в воде Черного моря - 1,3 раза.
Таким образом, сроки перехода в НС холерных вибрионов увеличивало как повышение температуры содержания микрокосм (1,6 -7,4 раза), так и искусственная их освещенность (1,3- 2,8 раза), что в сочетанном их влиянии оказывалось еще более выраженным (2,0 - 9,8 раза).
Зависимость образования НФ холерных вибрионов от солености сред, выразилась в более продолжительных (1,8 - 3,7 раза) сроках формирования НФ холерных вибрионов как при 4-6 0С без освещения во всех средах, содержащих микроэлементы, так и при 8-10 0С (2,8 - 6,4 раза). Сочетанное влияние среды микрокосм с повышением температуры их содержания и с искусственной освещенностью еще более (7,7 раза) увеличило сроки формирования НФ холерных вибрионов.
Наиболее продолжительным оказался период перехода в НС в микрокосмах с водой из Черного моря, характеризующейся наибольшим по сравнению со средами других микрокосм, даже водой Азовского моря, содержанием солей. Очевидно, сложный или более разнообразный минеральный состав воды Черного моря по сравнению с другими экспериментальными экосистемами способствует более продолжительному сохранению холерных вибрионов в культивируемом состоянии, имеющих в основном прототрофный тип питания, что может косвенно свидетельствовать о значении минерализации среды в поддержании культивируемого состояния вибрионов.
На скорость образования НФ холерных вибрионов в дистиллированной воде не влияли ни освещенность, ни температура содержания, что, очевидно, связано с минимальным содержанием в этой среде солей и увеличении питательного субстрата только в результате гибели и распада вибрионов.
Таким образом, низкие температуры, являющиеся основным фактором, запускающим переход холерных вибрионов в НС, в зависимости от состава экосистем реализуют индуцирующее действие в различные сроки. В дистиллированной воде, не содержащей солей и других факторов, влияющих на продолжительность сохранения холерных вибрионов в культивируемом состоянии, повышение температуры содержания микрокосм с 4-6 0С до 8-10 0С не удлиняло сроки перехода их в НС. Оценивая значимость экспериментальных данных о влиянии температуры на переход холерных вибрионов в НС, следует учитывать, что зимой с понижением температуры поверхностные холодные воды с температурой ниже 4 oС располагаются над сравнительно теплыми. По всей вероятности среда с температурой 4-5 oС является пограничной зоной в формировании некультивируемых форм у микроорганизмов, что коррелирует с получением экспериментальных НФ именно при этой температуре (5,6).
Список литературы
- Бароян О.В., Бургасов П.Н., Гайлонская И.Н., Мединский Г.М. Экология холерных вибрионов. //Вест. - АМН. - 1975, - №2, - С.45-53.
- Литвин В.Ю. Холера как природно-очаговая сапронозная инфекция //Журн. микробиол, эпидемиол. и иммунобиол. - 1995. - №6 - С. 30-31.
- Подосинникова Л.С., Соколенко А.В., Чепкова Е.А. и др. Экспериментальное получение и характеристика некультивируемых форм холерных вибрионов //Пробл. Комиссия «Холера и патоген. для человека вибрионы. Ростов н/Д, 1999. Вып. 12. - С. 13-14.
- Романова Ю.М., Чегаева Е.В., Гинцбург А.Л. Некультивируемое состояние у патогенных бактерий: известные и возможные факторы индукции обратимого процесса. //Мол. генет., микробиол. и вирусол. - 1998. - №3. - С.3-8.
- Савельев В.Н., Грижебовский Г.Н., Брюханов А.Ф. Некультивируемые формы холерного вибриона и их эпидемиологическое значение //Пробл. комиссия «Холера и патоген. для человека вибрионы». Ростов н/Д, 2001.- №14. - С.24-26.
- Соколенко А.В. Морфология, ультраструктура, метоболизм некультивируемых форм холерных вибрионов.: Автореф. дис....канд. биол. наук. -Ростов н/Д., 2000.- 18с.
- Colwell R.R., Brayton P.R., Grimes D. I. et al. Viable but non-culturable Vibrio cholerae and related pathogens in the environment: implication for release of genetically engineered microorganisms //BioTechnology.- 1985. - Vol.3. - P.817-820.
- Oliver J.D., Bockian R. In vivio resuscitation and virulence towerdes mice, of viable but nonculturable cells of Vibrio vulnificus //Appl. Envirion. Microbiol. - 1995. - Vol.61. - №7. - P. 2620-2627.
- Roszak D.B., Colwell R.B. Survival strategies of bacteria in the natural environment //Microbiol. Rev.-1987.-Vol.51.№3. - P.365-379.
Библиографическая ссылка
Николеишвили Л.Р, Подосинникова Л.С ПЕРЕХОД ХОЛЕРНЫХ ВИБРИОНОВ В НЕКУЛЬТИВИРУЕМОЕ СОСТОЯНИЕ ПОД ВЛИЯНИЕМ НЕКОТОРЫХ АБИОТИЧЕСКИХ ФАКТОРОВ // Фундаментальные исследования. 2004. № 2. С. 18-21;URL: https://fundamental-research.ru/en/article/view?id=2797 (дата обращения: 01.07.2026).