


Scientific journal
Fundamental research
ISSN 1812-7339
"Перечень" ВАК
ИФ РИНЦ = 1,798
THE PREGENERATIVE PERIOD ONTOGENY OF SOME SPECIES OF SUBFAMILY SEDOIDEAE BERGER IN CONDITIONS OF THE BELGOROD AREA

В последние годы значительно возрос интерес к ландшафтному дизайну. Озеленение улучшает эстетический облик городов и способствует снижению уровня влияния неблагоприятных факторов среды. Представители подсемейства Sedoideae Berger очень широко используются в озеленении благодаря своей декоративности и неприхотливости. Они применяются в создании альпийских горок, рокариев, рабаток, бордюров, фоновых куртин, в цветочных картинах, клумбах и различных композициях [2].
Изучение особенностей онтогенеза растений важно в интродукционных исследованиях, позволяет прогнозировать состояние растений через определенные промежутки времени, разрабатывать агротехнические мероприятия по уходу за насаждениями. Такие исследования отражают различные связи в области морфогенеза растений (между образованием корней, побегов, листьев, партикуляцей и другими процессами) и дают возможность понять причины их старения и отмирания [1].
Изучение прегенеративного периода отражает интенсивность нарастания и сроки взросления полезных растений. Исследование родовых комплексов позволяет сделать обоснованный отбор видов и сортов, наиболее устойчивых в культуре в местных условиях и перспективных для широкого использования в озеленении.
Цель исследований - выявить особенности онтогенетических состояний прегенеративного периода онтогенеза очитковых в условиях Белгородской области.
Материалы и методы исследования
Объектами изучения были виды, подвиды, разновидности, формы и сорта трех родов - Sedum L., Phedimus Raf., Hylotelephium H. Ohba - подсемейства Sedoideae (Ph. spurius (M. Bieb.) ‘t Hart, Ph. spurius1, Ph. spurius2, Ph. spurius3 (цифрами 1, 2, 3 обозначены сорта неизвестного происхождения), Ph. spurius var. variegatum, Ph. spurius ‘Album´, Phedimus kamschaticus (Fisch) ‘t Hart, Ph. kamschaticus f. variegatum, Ph. kamschaticus f. album, Ph. hybridus (L.) ‘t Hart, Ph. aizoon L., S. hispanicum L., S. acre L., S. sexangulare L., H. telephium subsp. maximum (L.) H. Ohba, H. cauticola (Praeger) H. Ohba.), перспективные для озеленения южных регионов России [8], интродуцированные в ботсад Белгородского государственного университета.
Семена вышеперечисленных видов и сортов, собранные в июле-августе, проращивали в феврале-марте 2009-2010 гг. в лабораторных условиях. Посев производили в плошки. Субстрат готовили из равных частей песка и почвы, предварительно простерилизованных. Семена проращивали при температуре 25-30 °С, затем полученные всходы содержали в комнатных условиях при температуре 22-24 °С. После достижения сеянцами виргинильного онтогенетического состояния была произведена их высадка в открытый грунт.
Определение онтогенетических состояний особей проводили на основании комплекса качественных морфологических и биологических признаков с использованием методических подходов Т.А. Работнова, А.А. Уранова, авторских коллективов и А.В. Поповой [3-7]. В течение периода развития сеянцев проводили наблюдения за состоянием растений, устанавливали сроки наступления оногенетических состояний, описывали их морфологические характеристики.
Результаты исследований и их обсуждения
Изученные представители очитковых характеризуются быстрой всхожестью семян при соблюдении оптимального температурно-влажностного режима.
Проростки у всех исследованных представителей очитковых сходны. Они имеют два семядольных листика светло-зеленой окраски, тонкие, цельнокрайние, неопушенные, широкоовальной формы, диаметром 0,5-1 мм. У H. telephium subsp. maximum семядольные листики были крупнее - до 2 мм. Гипокотиль тонкий, длиной 0,3-1,2 см, от белой до бледно-зеленой окраски. Длина главного корня 1-2 мм, у большинства проростков развивались боковые корни первого порядка (табл. 1).
Таблица 1
Морфологические схемы сеянцев в разных онтогенетических состояниях
|
Вид, подвид |
Онтогенетические состояния |
|||
|
p |
j |
im |
v |
|
|
Hylotelephium telephium |
|
|
|
|
|
Phedimus spurius, |
|
|
|
|
|
Sedum acre, |
|
|
|
|


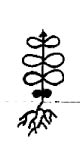
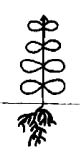



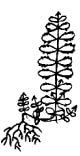




Примечание: заштрихованные элементы - листики семядолей.
Продолжительность состояния проростка в среднем составляла от 10 до 20 дней, больше 20 дней отмечено у Ph. hybridus (до 25 дней) и H. telephium subsp. maximum (до 28 дней).
Переход в ювенильное онтогенетическое состояние у изученных видов происходит на 12-15 день. У Ph. spurius2, Ph. kamtschaticus f. variegatum и H. telephium subsp. maximum ювенильное состояние наступало позже (табл. 2). Меньше по времени в данном состоянии находился вид H. cauticola (12-15 дней), дольше - Ph. aizoon (58-62 дня), Ph. hybridus (84-89 дней). Признаком перехода в ювенильное состояние служило образование первой пары настоящих листьев. Отмирание семядольных листиков в данном случае не являлось признаком перехода (см. табл. 2). Побеги очитковых характеризовались небольшими размерами - длиной в среднем 1,0-3,5 см, самые мелкие сеянцы были у H. cauticola (до 1 см). Длина эпикотиля - от 0,2 до 0,7 см; у S. acre, S. sexangulare, H. cauticola эпикотиль не заметен. Сеянцы видов рода Phedimus и вида H. cauticola имели по 2-4 метамера и два листа в узле. Виды рода Sedum имели наибольшее число метамеров - до 12 и очень короткие междоузлия. У H. telephium subsp. maximum отмечено наименьшее число метамеров - 1-2. Листорасположение у Ph. hybridus очередное, у всех остальных сеянцев - супротивное. У вида S. hispanicum первая и вторая пара настоящих листьев располагаются супротивно, с третьего узла - мутовчатое: по четыре листа в мутовке и в дальнейшем развивается по четыре настоящих листа одновременно.
Таблица 2
Развитие сеянцев очитковых в лабораторных условиях
|
Название |
Появление 1-х всходов, день |
Массовое прорастание семян, день |
День начала формирования некоторых элементов главного побега от появления всходов в различных онтогенетических состояниях* |
Наступление виргинильного состояния (v), день |
|||||||
|
p |
j |
im |
|||||||||
|
Корни |
1-я пара листьев |
2-я пара листьев |
3-я пара листьев |
4-я пара листьев |
Первые боковые побеги |
Корни |
Корни |
||||
|
Ph. spurius |
3-й |
5-й |
10-й |
13-й |
22-й |
29-й |
36-й |
51-й |
25-й |
31-й |
86-й |
|
Ph. spurius1 |
2-й |
5-й |
9-й |
16-й |
26-й |
33-й |
39-й |
56-й |
36-й |
56-й |
89-й |
|
Ph. spurius2 |
5-й |
12-й |
10-й |
20-й |
40-й |
51-й |
61-й |
66-й |
51-й |
73-й |
98-й |
|
Ph. spurius3 |
3-й |
8-й |
13-й |
12-й |
21-й |
32-й |
40-й |
50-й |
40-й |
48-й |
94-й |
|
Ph. spurius var. variegatum |
2-й |
5-й |
10-й |
13-й |
22-й |
34-й |
46-й |
57-й |
27-й |
34-й |
86-й |
|
Ph. spurius Album |
2-й |
4-й |
9-й |
15-й |
21-й |
32-й |
39-й |
76-й |
31-й |
51-й |
101-й |
|
Ph. kamtschaticus |
3-й |
4-й |
8-й |
12-й |
22-й |
35-й |
47-й |
62-й |
46-й |
60-й |
97-й |
|
Ph. kamtschaticus f. variegatum |
3-й |
5-й |
12-й |
44-й |
52-й |
64-й |
73-й |
65-й |
67-й |
105-й |
125-й |
|
Ph. kamtschaticus f. album |
2-й |
4-й |
13-й |
13-й |
22-й |
31-й |
36-й |
66-й |
26-й |
33-й |
86-й |
|
Ph. hybridus |
3-й |
4-й |
10-й |
21-й |
32-й |
67-й |
98-й |
105-й |
26-й |
69-й |
120-й |
|
Ph. aizoon |
3-й |
3-й |
10-й |
13-й |
30-й |
41-й |
57-й |
71-й |
41-й |
59-й |
90-й |
|
S. acre |
2-й |
6-й |
13-й |
14-й |
22-й |
33-й |
36-й |
61-й |
59-й |
89-й |
96-й |
|
S. sexangulare |
3-й |
5-й |
19-й |
14-й |
22-й |
34-й |
36-й |
50-й |
33-й |
59-й |
100-й |
|
S. hispanicum |
2-й |
4-й |
11-й |
11-й |
14-й |
21-й |
26-й |
30-й |
27-й |
33-й |
80-й |
|
H. telephium subsp. maximum |
2-й |
5-й |
11-й |
25-й |
45-й |
62-й |
68-й |
- |
51-й |
64-й |
175-й |
|
H. cauticola |
3-й |
5-й |
13-й |
13-й |
27-й |
37-й |
51-й |
- |
- |
- |
- |
Примечания: * p - проросток, j - ювенильное онтогенетическое состояние, im - имматурное онтогенетическое состояние.
Форма листьев от обратно-широкояйцевидной (вид Ph. spurius, его разновидности и сорта) до обратнояйцевидной (вид Ph. kamtschaticus и его разновидности, вид Ph. hybridus), широкояйцевидная - у Ph. aizoon, H. telephium subsp. maximum, H. cauticola. У видов рода Sedum форма листьев варьирует: у S. hispanicum - удлиненно-ланцетная, S. acre - яйцевидная, S. sexangulare - игольчатая.
Для представителей рода Phedimus характерно наличие зубцов с каждой стороны листовой пластинки. Со второй или третей пар листьев начинают проявляться один или два зубца. С третьей или четвертой пары листьев число зубцов увеличивается. Максимальное число зубцов - четыре, у Ph. kamtschaticus - пять, Ph. aizoon - восемь.
Размеры листьев у большинства очитковых были сходны - длиной 0,5-1,0 см и шириной 0,3-0,5 см. Наиболее крупные листья имели сеянцы Ph. aizoon - длиной до 1,5 см и шириной до 1 см. Самые мелкие листья характерны для представителей рода Sedum - длиной от 0,2 до 0,5 см, шириной 0,1-0,2 см.
Имматурное онтогенетическое состояние наступало раньше у сеянцев S. hispanicum - на 30 день после появления всходов, позже - у Ph. hybridus (на 105 день) и H. telephium subsp. maximum (на 175 день), у остальных исследованных очитковых - в среднем на 50-70 день. Наименьшая продолжительность нахождения растений в данном состоянии была у Ph. hybridus и Ph. aizoon (15-22 дня), наибольшая продолжительность - у H. telephium subsp. maximum (124-127 дней). У вида H. cauticola имматурное состояние не отмечено. Сеянцы очень слабо росли, частично погибали. Для дальнейшего их развития, возможно, требуются специфические условия. У остальных видов очитковых продолжительность имматурного состояния в среднем составляла 30-60 дней. Признаком перехода в имматурное состояние было образование первых боковых побегов в пазухах семядольных листьев, развитие боковых корней второго и третьего порядков, начало полегания сеянцев (см. табл. 1).
Сеянцы характеризовались более крупными размерами, высотой побегов 6-10 см. Наименьшая высота побегов отмечены у S. acre и S. sexangulare - 2,0-4,5 см, но с наибольшим числом метамеров (8-18). Наименьшее число метамеров было у H. telephium subsp. maximum как и в ювенильном состоянии. У остальных сеянцев в среднем число метамеров составляло 4-7. Форма листьев и листорасположение не изменялось, за исключением S. acre, у которого листорасположение сменилось на очередное. Листья у большинства очитковых длиной 1-2,5 см и шириной 0,5-1,7 см. Наиболее крупные листья формируются у Ph. aizoon (длиной до 3 см и шириной до 1,7 см) как и в более ранних состояниях, мелкими листьями характеризуются представители рода Sedum - длиной от 0,2 до 0,7 см, шириной 0,1-0,3 см.
Переход в виргинильное онтогенетическое состояние происходит на 80-125 день после появления всходов, позже у H. telephium subsp. maximum (на 183 день). Наименьшая продолжительность этого состояния отмечена для H. telephium subsp. maximum (8-12 дней) и Ph. aizoon (23-25 дней). Эти виды очитковых в первый год жизни достигают молодого генеративного состояния. У других изученных представителей рода Phedimus виргинильное онтогенетическое состояние длится 1-2 года, у видов Sedum - от одного до трех лет. Качественными признаками перехода в данное состояние является появление основных черт, характерных для взрослых растений - дефинитивные листья, ветвящиеся побеги и корни (см. табл. 1).
В виргинильном состоянии побеги достигают в среднем 10-20 см, самые низкие сеянцы были у S. acre и S. sexangulare, S. hispanicum, H. telephium subsp. maximum - высотой 3-9 см. Число метамеров у видов рода Phedimus и S. hispanicum - 9-13. Наибольшим числом метамеров характеризуются S. acre и S. sexangulare (10-35), наименьшим - H. telephium subsp. maximum (3-4). Форма листьев и листорасположение у виргинильных растений не меняется. В среднем у большинства очитковых листья достигают 1-3 см в длину и 0,6-2,0 см в ширину. У вида Ph. aizoon с самыми крупными листьями параметры увеличиваются до 4 см в длину и 2,5 см в ширину, у мелколистных видов рода Sedum до 0,3-1,0 см в длину, 0,1-0,4 см в ширину.
Корневая система представлена морфологически однородными ветвящимися корнями. Главный корень не выделяется среди корней различного происхождения. В данном состоянии сеянцы легко адаптируются к условиям открытого грунта и благополучно переносят пересадку.
Заключение
В результате изучения онтогенетических состояний прегенеративного периода очитковых выявлены количественные параметры, характерные для различных онтогенетических состояний - число метамеров, размеры листьев, высота побегов. Выявлены различия сроков онтогенетических состояний (начало и продолжительность) и качественные признаки для них - листорасположение, форма листьев, характер нарастания.
Обнаружены сходства у всех проростков изученных очитковых; у большинства из них образуются боковые корни. Вид S. hispanicum отличался самым коротким прегенеративным периодом и всех его онтогенетических состояний. Самые поздние сроки переходов из одного в другие онтогенетические состояния характерны для H. telephium subsp. maximum. Форма и листорасположение у большинства изученных видов (за исключением S. acre) были стабильными на протяжении прегенеративного периода. Самые крупные сеянцы развиваются у Ph. aizoon, самые мелкие - у видов рода Sedum, но они обладают наибольшим числом метамеров. Наименьшая продолжительность виргинильного состояния отмечена у H. telephium subsp. maximum (8-12 дней) и Ph. aizoon (23-25 дней). Данные виды в первый год жизни достигали молодого генеративного состояния.
Работа выполнена в рамках ФЦП «Научные и научно-педагогические кадры инновационной России» на 2009-2013 годы (госконтракт № 14.740.11.0739 от 12 октября 2010 г.).
Список литературы
- Игнатьева И.П. Онтогенетический морфогенез вегетативных органов травянистых растений: учебное пособие. - М.: Московская сельхоз. академия им. К.А. Тимирязева, 1983. - 53 с.
- Клюйков Е.В., Госсе Д.Д. Очитки и очитники. - М.: Изд. Дом МСП, 2006. - 48 с.
- Попова А.В. Начальные стадии онтогенеза некоторых видов рода Sedum L. (Crassulaceae DC.) // Проблеми збереження, вiдновлення та збагачення бiорiзномантностi в умовах антропогенного змiненого середовища: матерiали мiжнародноï науковi конференцiï. - Кривий Рiг, 2005. - С. 287-288.
- Работнов Т.А. Жизненный цикл многолетних травянистых растений в луговых ценозах. - М.- Л.: Тр. БИН АН СССР. - 1950. - Сер. 3, № 6. - С. 7-204.
- Уранов А.А. Возрастной спектр фитоценопопуляций как функция времени и энергетических волновых процессов // Биологические науки. - 1975. - №2. - С. 7-34.
- Ценопопуляции растений (основные понятия и структура) / Л.И. Воронцова, Л.Е. Гатцук, В.Н. Егорова и др. - М.: Наука, 1967. - 214 с.
- Ценопопуляции растений: (Очерки популяционной биологии) / Л.Б. Заугольнова, Л.А. Жукова, А.С. Комаров и др. - М.: Наука, 1988. - 184 с.
- Genera of Crassulaceae subfam. Sedoideae / United States Department of Agriculture. Germplasm Resources Information Network (GRIN). - 2007. - Режим доступа: http://www.ars-grin.gov/cgi-bin/npgs/html/gnlist.pl?1764.
Рецензенты:
Ткаченко И.К., д.с.-х.н., профессор, профессор кафедры анатомии и физиологии живых организмов Биолого-химического факультета Белгородского государственного университета Министерства образования и науки РФ, г. Белгород;
Кочкаров В.И., д.б.н., профессор кафедры фармхимии и фармакогнозии Фармацевтического факультета Белгородского государственного университета Министерства образования и науки РФ, г. Белгород.
Работа поступила в редакцию 09.03.2011.
Библиографическая ссылка
Орлова О.Н, Сорокопудова О.А ПРЕГЕНЕРАТИВНЫЙ ПЕРИОД ОНТОГЕНЕЗА НЕКОТОРЫХ ВИДОВ ПОДСЕМЕЙСТВА SEDOIDEAE BERGER В УСЛОВИЯХ БЕЛГОРОДСКОЙ ОБЛАСТИ // Фундаментальные исследования. 2011. № 8-1. С. 93-97;URL: https://fundamental-research.ru/en/article/view?id=26791 (дата обращения: 01.08.2026).