


Scientific journal
Fundamental research
ISSN 1812-7339
"Перечень" ВАК
ИФ РИНЦ = 1,798
INTRAMURAL LYMPH NODE AS VARIENT OF LYMPHATIC SYSTEM EVOLUTION

Введение
Лимфатические узлы (ЛУ) являются частью непрерывного экстраорганного лимфатического русла, обычно исследуются как иммунные органы [1,4]. В последнее время получила широкое распространение идея, что лимфатическая система в качестве самостоятельной части организма не существует: лимфатические сосуды (ЛС) обслуживают ЛУ и являются придатком лимфоидной системы [3]. Лимфатические пути образуются в эволюции и онтогенезе как выключенные из кровотока коллатерали венозного русла [2]. Сначала лимфатическая система представлена сетью ЛС. Позднее на стенках ЛС конденсируется лимфоидная ткань. Первые ЛС регистрируются у хрящевых рыб. В стенках грудного протока и других ЛС рептилий появляются лимфоидные узелки. У змей они вдаются в просвет лимфатических синусов, которые окружают аорту, полую и внутреннюю яремную вены. Первые ЛУ возникают у водоплавающих птиц, имеют нетипичное (для млекопитающих), инвертированное строение: их центральный синус соединяет афферентный и эфферентный ЛС и окружен корковым веществом с лимфоидными узелками, кнаружи расходятся добавочные, боковые синусы и мозговые тяжи. ЛУ птицы выглядит как слабое циркулярное впячивание растущей лимфоидной муфты ЛС (утолщение стенки с лимфоидной тканью) в его просвет, который сужается (центральный синус) и ветвится (боковые синусы). ЛУ клоачных млекопитающих имеет вид вторичного лимфоидного узелка в просвете ЛС (определяется широкий краевой синус). У плацентарных млекопитающих закладка ЛУ формируется как первичный периваскулярный лимфоидный узелок в просвете ЛС (рис. 1, 2).
Материалы и методы
Работа проведена на 30 белых крысах, усыпленных парами хлороформа. Были изготовлены: 1) серийные гистологические срезы грудного протока, окрашенные пикрофуксином по Ван Гизон; 2) тотальные препараты грудного протока и подвздошного лимфатического коллектора вместе с капсулой его пристеночного лимфоузла, окрашенные галлоцианином по Эйнарсону.
Результаты и их обсуждение
У белой крысы диффузная лимфоидная ткань постоянно встречается на протяжении грудного протока в виде пристеночных и внутристеночных скоплений лимфоцитов, в т.ч. в основании клапанных заслонок и в составе интралюминарных перегородок, которые на гистологических срезах напоминают клапанные створки. По таким перегородкам могут проходить только прободающие кровеносные сосуды. Подобные перегородки встречаются в лимфатических коллекторах у рыб и амфибий. Недавно я обнаружил внутристеночный ЛУ округлой формы, небольших размеров. Его капсула переходила в латеральную стенку клапанного синуса крупного подвздошного ЛС белой крысы (рис. 3). Эфферентные ЛС у этого ЛУ отсутствовали, воротный синус открывался прямо в клапанный синус данного ЛС, мелкие афферентные ЛС – в краевой синус ЛУ. На уровне его дистального края в ЛС находился двухстворчатый клапан. Одна из его комиссур срасталась с капсулой ЛУ – стенкой воротного синуса. Из ее толщи (из капсулярной комиссуры клапана) выходил веерообразный пучок гладких миоцитов. Проксимальнее клапана, в мышечной манжетке лимфангиона он сливался с другим комиссуральным пучком. Объединенный косопродольный пучок миоцитов охватывала поперечная мышечная петля. При сокращении подвздошного ЛС пучок может подтягивать в проксимальном направлении его клапан и внутристеночный ЛУ и способствовать его опорожнению. Несмотря на необычность строения такого ЛУ, его морфогенез можно представить по стандартной схеме: кровеносные сосуды инвагинируют в расширяющийся просвет ЛС вместе с его эндотелиальной стенкой, межсосудистая соединительная ткань преобразуется в лимфоидную вокруг кровеносных микрососудов инвагинации. Обычно наиболее крупные vasa vasorum (артериола и венула) ЛС определяются в основании его клапанов. Подвздошный лимфатический коллектор лежит сбоку от одноименных кровеносных сосудов. Их ветви могут пересекать ЛС, инвагинировать в устье притока его клапанного синуса и обусловить закладку внутристеночного ЛУ. При такой локализации ЛУ нет необходимости в эфферентном ЛС. Обычно в этой области находятся крупный подвздошный или каудальный поясничный ЛУ эллипсовидной формы. Их ворота с кровеносными сосудами и эфферентными ЛС обращены к подвздошным сосудам и бифуркации аорты.
 .
.
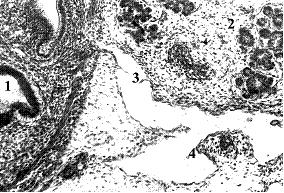
Рис. 1. Плод человека 10,5 нед., поперечный срез: 1 – стенка двенадцатиперстной кишки; 2 – головка поджелудочной железы; 3 – двухстворчатый клапан заднего поджелудочно-двенадцатиперстного лимфатического сосуда; 4 – стромальный зачаток панкреато-дуоденального лимфатического узла с артериолой и венулой. Гематоксилин и эозин. Ув.120
 .
.
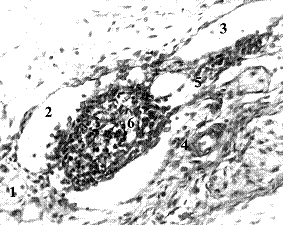
Рис. 2. Плод человека 13 нед., поперечный срез: 1,3 – афферентный и эфферентный лимфатические сосуды панкреато-дуоденального лимфоузла; 2 – его краевой синус, намечается тонкая капсула; 4,5 – артериола и венула в формирующихся воротах узла; 6 – недифференцированная паренхима узла. Рост лимфоидного зачатка в просвете матричного лимфатического сосуда приводит к расширению этого участка сосуда сверх ширины его смежных участков. Гематоксилин и эозин. Ув. 300.

 .
.
А Б
Рис. 3. Внутристеночный подвздошный лимфатический узел крысы, тотальный препарат капсулы. А: 1 – афферентные лимфатические сосуды, их стенки переходят в капсулу узла; 2 – стенка воротного синуса; 3,4 – двухстворчатый клапан подвздошного лимфатического сосуда; 5 – комиссуральный пучок миоцитов. Б: 2 – стенка воротного синуса; 3,4 – поперечные, дугообразные пучки миоцитов в клапанных валиках; 5 – комиссуральный пучок миоцитов; 6 – веерообразный пучок миоцитов, который выходит из сращения клапана с капсулой узла; 7 – поперечная мышечная петля, она охватывает срастающиеся косопродольные пучки (5) и (6); 8,9 – дистальные клапанные пучки миоцитов, в т.ч. прободающие клапанный валик. Галлоцианин. Ув.: А – 100; Б – 300.
Заключение
Как и в эмбриогенезе, в эволюции поначалу (у рыб и амфибий) кровеносные сосуды пересекают ЛС и с окружающей соединительной тканью инвагинируют в их просвет (стромальные зачатки ЛУ, или предузлы). Лимфоидная ткань в стенках ЛС (интрамуральная) появляется позднее (у рептилий и птиц) и становится основой морфогенеза ЛУ у водоплавающих птиц в виде лимфоидной муфты вокруг центрального синуса ЛУ. У млекопитающих интравазальная (погруженная в просвет ЛС) лимфоидная ткань в виде лимфоидного узелка с краевым синусом становится ЛУ. У крысы я нашел промежуточное по строению образование – внутристеночный ЛУ. Он имеет вид варикозного выроста клапанного синуса ЛС с лимфоидной тканью и краевым синусом в стенке и может служить аргументом в пользу эволюционной трансформации пристеночных скоплений лимфоидной ткани ЛС в ЛУ. Переход от внутристеночного лимфоидного предузелка ЛС к типичному ЛУ обусловлен интенсификацией деформации ЛС кровеносными сосудами под давлением растущих органов. Ранее [2] я уже обращал внимание на корреляцию вправления физиологической пупочной грыжи в брюшную полость плода человека и крысы с закладкой поясничных и брыжеечных лимфоузлов. У птиц вправление грыжи происходит на более поздних стадиях эмбриогенеза (печень гораздо меньше), а у рептилий – после вылупления детеныша из яйца, когда ЛС уже приобрели наружную оболочку. Можно предположить, что нарастающее в эволюции позвоночных внешнее давление органов и кровеносных сосудов на стенки ЛС играет важную роль в возникновении ЛУ. В эволюции позвоночных животных наблюдается интенсификация их основного обмена веществ, а в этой связи – процессов роста органов и гистогенеза, в т.ч. иммуногенеза. Поэтому не случайно ЛУ, с учетом механики их морфогенеза, появляются у теплокровных животных.
СПИСОК ЛИТЕРАТУРЫ
1. Бородин Ю.И., Сапин М.Р., Этинген Л.Е. и др. (ред.) Функциональная анатомия лимфатического узла. – Новосибирск: Наука, 1992. – 257 с.
2. Петренко В.М. Эволюция и онтогенез лимфатической системы. – СПб.: СПбГМА, ДЕАН, 2003. – 336 с.
3. Сапин М.Р. // Морфология. – 1998. – Т.113, № 3. – С. 106.
4. Сапин М.Р., Этинген Л.Е. Иммунная система человека. – М.: Медицина, 1996. – 408 с.
Библиографическая ссылка
В.М. Петренко ВНУТРИСТЕНОЧНЫЙ ЛИМФАТИЧЕСКИЙ УЗЕЛ КАК ВАРИАНТ ЭВОЛЮЦИИ ЛИМФАТИЧЕСКОЙ СИСТЕМЫ // Фундаментальные исследования. 2009. № 8. С. 39-42;URL: https://fundamental-research.ru/en/article/view?id=1982 (дата обращения: 30.06.2026).