



Введение
Белая крыса является важным экспериментальным животным, на котором изучают, в частности, механизмы лимфотока. Однако его структурные основы в лимфатических узлах (ЛУ), в т.ч. брыжеечных, и развитие ЛУ крысы мало изучены. У человека гораздо больше ЛУ разных групп и самих групп ЛУ, в т.ч. верхних (краниальных) брыжеечных ЛУ, чем у крысы [1,3-5]. В развитии ЛУ можно выделить три стадии - закладки (I фаза - стромальных зачатков, II фаза - лимфоидных зачатков), первичной и вторичной дифференциации [2,3]. Закладка верхних брыжеечных ЛУ происходит у плодов человека 3-го мес. путем инвагинации верхних брыжеечных артерии и вены с их ветвями и притоками, эндотелиальной стенкой сопровождающих лимфатических сосудов (ЛС) и межсосудистой рыхлой соединительной тканью в расширяющийся просвет ЛС. Так возникают стромальные зачатки брыжеечных ЛУ. Уже у плодов 11-12 нед. макрофаги и лимфоциты мигрируют в строму инвагинаций из кровеносных микрососудов. В результате стромальные зачатки превращаются в лимфоидные. Их рост сопровождается локальным сужением и искривлением просвета матричного ЛС с образованием первичного краевого синуса ЛУ. «Ножки» инвагинаций (пристеночные основания зачатков ЛУ) сужаются и преобразуются в ворота ЛУ. Матричный ЛС разделяется на первичный краевой синус, афферентные и эфферентные ЛС брыжеечного(ых) ЛУ. Первичная дифференциация (плоды 4-го мес.) проявляется обособлением ЛУ - тонкая капсула с густой сетью ретикулярных волокон, краевой синус, отделенный от ЛС клапанами, воротный синус как локальное расширение краевого синуса. Вторичная дифференциация верхних брыжеечных ЛУ начинается у плодов 5-го мес. Наблюдаются ветвление краевого синуса ЛУ, образование и рост его промежуточных синусов, расширение воротного синуса, принимающего промежуточные синусы, хиларное утолщение капсулы и рост трабекул, дифференциация вещества ЛУ, зональная (корковое и мозговое) и локальная (лимфоидные узелки). ЛУ приобретают первичные узелки у плодов 5-7 мес., вторичные - обычно после рождения. У плодов 4-5 мес. капсула ЛУ, как и стенка сопряженных ЛС, разделяется на слои - внутренний, с густой сетью тонких ретикулярных волокон, и наружный, с более толстыми ретикулярными и первыми коллагеновыми волокнами. На границе между ними появляются первые миоциты. Во второй половине утробной жизни человека наблюдается рост пучков гладких миоцитов и эластических волокон в капсулах и трабекулах ЛУ. Появляются дополнительные мышечные слои в капсуле ЛУ - постоянный, но очень рыхлый в наружном слое и непостоянный, но с лучше выраженными мышечными пучками во внутреннем слое [2,3].
Материал и методы
Работа выполнена на 30 белых крысах обоего пола 7-30 сут., 20 плодах белой крысы 16-21 сут. и 5 новорожденных крысятах (1-е сутки жизни). Крыс усыпляли хороформом, затем фиксировали в жидкости Буэна. Изготавливали серии поперечных срезов плодов и новорожденных крысят и срезов органокомплексов с корнем брыжейки тонкой кишки крыс 7-30 сут. толщиной 5-7 мкм. Срезы окрашивали азур-II-эозином, гематоксилином и эозином, смесью Маллори, пикрофуксином по Ван Гизон, импрегнировали азотнокислым серебром по Футу.
Результаты
Закладка брыжеечных ЛУ происходит у плодов крысы 17-18 сут. В корне брыжейки тонкой кишки и около двенадцатиперстно-тощекишечного изгиба залегают краниальные брыжеечные артерия и вена, а по обе стороны от них - крупные ЛС с эндотелиальными стенками. Артерия и ее ветви, вена и ее притоки обладают более толстыми и дифференцированными стенками, которые включают наружную оболочку. Они вместе с эндотелием ЛС и межсосудистой рыхлой соединительной тканью инвагинируют в просвет ЛС и разделяют его на первичные краевые синусы стромальных зачатков ЛУ (инвагинация) и их ЛС, афферентные и эфферентные. Закладки брыжеечных ЛУ полиморфны, различаются прежде всего длиной и числом ворот (места инвагинаций ветвей артерии и притоков вены). У плодов крысы 18-19 сут. межсосудистая рыхлая соединительная ткань инвагинаций преобразуется в лимфоидную, а стромальные зачатки ЛУ - в лимфоидные. Первичная дифференциация брыжеечных ЛУ (плоды крысы 19-20 сут.) характеризуется их обособлением: растущий зачаток все более преобладает над матричным ЛС по диаметру, что сопровождается удлинением, сужением и искривлением краевого синуса ЛУ. Его наружная эндотелиальная стенка дополняется очень тонким слоем соединительной ткани - образуется тонкая капсула, которая содержит все более густую сеть ретикулярных волокон. Краевой синус отделяется от ЛС клапанами, намечается воротный синус как небольшое расширение краевого синуса около «ножки» инвагинации, которая становится воротами ЛУ. В его гомогенную лимфоидную паренхиму врастают ветви внутренней стенки краевого (подкапсульного) синуса ЛУ с образованием промежуточных синусов (рис.1). Их эндотелиальная выстилка прогрессивно истончается и разрежается. К ней непосредственно примыкают очень тонкие ветви капсулы (трабекулы) и лимфоидная ткань. Вторичная дифференциация брыжеечных ЛУ начинается перед рождением крысят (21-е сут.). Продожаются ветвление краевого и рост промежуточных синусов, расширение воротного синуса, хиларное утолщение капсулы и рост трабекул, капсулярных и хиларных. У новорожденных крысят капсула ЛУ, как и стенка ЛС, разделяется на слои - внутренний, с густой сетью тонких ретикулярных волокон, и наружный, с более толстыми ретикулярными и первыми коллагеновыми волокнами. На границе между ними появляются первые гладкие миоциты, их растущие пучки позднее проникают в трабекулы.
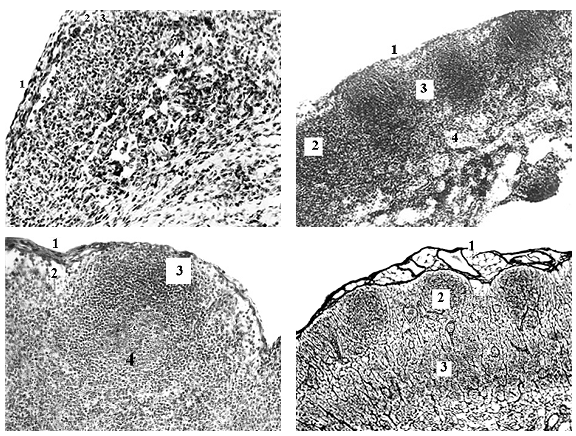
Рис. 1. Брыжеечный лимфатический узел крысы. А - плод 21 сут: 1 - капсула; 2 - краевой синус; 3,4 - корковое и мозговое вещество. Гематоксилин и эозин. Ув. 200. Б - плод 14 сут: 1 - капсула; 2 - первичный лимфоидный узелок; 3 - межузелковая лимфоидная ткань; 4 - мозговой тяж. Гематоксилин и эозин. Ув. 100. В - плод 21 сут: 1 - капсула; 2 - краевой синус; 3 - вторичный лимфоидный узелок и 4 - его герминативный центр. Азур-II-эозин. Ув. 300. Г - плод 28 сут: 1 - капсула; 2 - лимфоидный узелок; 3 - светлый центр Т-домена. Серебрение по Футу. Ув. 100.
Разделение вещества брыжеечных ЛУ белой крысы на корковое и мозговое начинается у плодов 21 сут, в присутствии посткапиллярных венул с высокими эндотелиоцитами. У новорожденных крысят (1-е сут. жизни) корково-мозговой индекс (К/М) этих ЛУ равен 3,5, расширяются их корковые синусы. Через неделю К/М-индекс ЛУ уменьшается до 3,0. Около краевого синуса появляются первые, единичные лимфоидные узелки без герминативных центров. На их месте наблюдается разрыхление сети ретикулярных волокон. В результате корковое вещество начинает разделяться на периферическую и глубокую зоны. В мозговых тяжах обнаружены плазмоциты, их доля среди лимфоидных клеток равна 8,5%. К 2-м нед жизни К/М-индекс брыжеечных ЛУ уменьшается до 2,55. Множественные (5,89±0,28 в поле зрения, ув.100) лимфоидные узелки не имеют герминативных центров. Напротив первичных лимфоидных узелков в глубокой коре ЛУ обнаружено разрыхление ретикулярной стромы, но размеры дифференцирующихся светлых центров Т-доменов меньше самих узелков. Доля плазмоцитов в мякотных тяжах достигает 20%. У крысят 3 нед К/М-индекс брыжеечных ЛУ немного уменьшается (2,33). Общее количество лимфоидных узелков достигает 9,88±0,44, у части из них (5,2±0,3; 52,63% от всех узелков) определяются герминативные центры. Расширяются светлые центры Т-доменов. Их ширина сопоставима с диаметром лимфоидных узелков. Доля плазмоцитов в клеточном составе мозговых тяжей увеличивается до 25%. У крыс 4 нед К/М-индекс брыжеечных ЛУ приближается к дефинитивному (2,0) значению. Общее количество лимфоидных узелков составляет 13,1±0,5, в т.ч. вторичных узелков - 7,9±0,4 (60,31% от всех узелков). Наблюдается слияние нескольких первичных очагов разрыхления стромы глубокой коры в единый для 3-4 лимфоидных узелков светлый центр Т-домена.
Заключение
Полученные данные показывают, что брыжеечные ЛУ белой крысы и человека проходят сходные стадии развития, но в различные сроки онтогенеза. Вторичная дифференциация ЛУ начинается в середине утробной жизни человека и завершается (появление вторичных лимфоидных узелков) после его рождения, а у белой крысы - начинается перед самым ее рождением и завершается (морфогенез лимфоидных узелков) после рождения крысы. Однако следует учитывать, что новорожденные крысята находятся на стадии развития, эквивалентной плодам человека 5-6 мес. Первичные лимфоидные узелки в ЛУ белой крысы дифференцируются в связи с усилением антигенной стимуляции после рождения (переход во внешнюю среду), вторичные узелки ЛУ - после перехода крысенка на смешанный тип питания (с 2 нед. жизни). Плазмоциты появляются первоначально в мозговых тяжах ЛУ, когда появляются первые лимфоидные узелки, задолго (за 2 нед.) до обнаружения в них герминативных центров. Источником пренатальной антигенной стимуляции могут служить продукты жизнедеятельности самого плода, в частности распада временных структур. Дифференциация ЛУ как лимфатического и лимфоидного органа ускоряется после рождения (число и размеры соединительнотканных волокон, миоцитов, лимфоцитов и плазмоцитов - превалирует «внутренний» рост ЛУ), что свидетельствует о значительном увеличении функциональной нагрузки.
СПИСОК ЛИТЕРАТУРЫ
- Жданов Д.А. Хирургическая анатомия грудного протока и главных лимфатических коллекторов и узлов туловища. - Горький: Изд-во Горьковск. мед.ин-та, 1945. - 308 с.
- Петренко В.М. Развитие лимфатической системы в пренатальном онтогенезе человека. - СПб. Изд-во СПбГМА, 1998. - 364 с.
- Петренко В.М. Эволюция и онтогенез лимфатической системы. - Изд 2-е. - СПб. - Изд-во ДЕАН, 2003. - 336с.
- Рахимов Я.А. Грудной проток млекопитающих. - Душанбе. Изд-во «Ирфон» 1968. - 214 с.
- Сапин М.Р., Борзяк Э.И. Внеорганные пути транспорта лимфы. - М.: Изд-во Медицина, 1982. - 264 с.
Библиографическая ссылка
В.М. Петренко, Е.В. Петренко РАЗВИТИЕ БРЫЖЕЕЧНЫХ ЛИМФАТИЧЕСКИХ УЗЛОВ У КРЫСЫ // Фундаментальные исследования. 2010. № 5. С. 7-11;URL: https://fundamental-research.ru/ru/article/view?id=6977 (дата обращения: 04.07.2026).