



Пусть 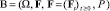 – стохастический базис с обычными условиями Деллашери [1]. Пусть при t ∈ [0, 1] заданы точечные процессы
– стохастический базис с обычными условиями Деллашери [1]. Пусть при t ∈ [0, 1] заданы точечные процессы 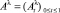 и
и 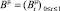 , определяющие позиции активных транспозонов на хромосомах с нормированными длинами (равными 1). В таких обозначениях наличие активных транспозонов может быть обозначено единичным значением процесса телеграфного типа
, определяющие позиции активных транспозонов на хромосомах с нормированными длинами (равными 1). В таких обозначениях наличие активных транспозонов может быть обозначено единичным значением процесса телеграфного типа
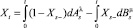 (1)
(1)
с процессами Aλ и Bμ, имеющими компенсаторы
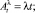
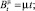
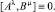 (2)
(2)
Обозначим xt = EXt. Тогда, очевидно, выполняется соотношение
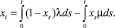 (3)
(3)
Обозначим среднюю длину транспозона β (β > 0).
Тогда очевидно
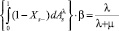 , (4)
, (4)
поскольку, как следует из уравнения (3),
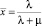 (5)
(5)
(при стационарном распределении, наблюдаемом в экспериментах x′ = 0, и, следовательно, выполняется (5)).
Из (4) вытекает полезное соотношение для соотношения частоты затухания активности транспозонов и их средней длины
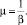 (6)
(6)
Пусть в клетке существует k ≥ 1 (идентичных) копий хромосомы. Тогда локусы активности транспозонов, разрушающие работу нормальной ДНК, могут быть обозначены 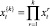 со средними значениями на всех k хромосомах, равными
со средними значениями на всех k хромосомах, равными
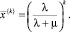 (7)
(7)
Последнее равенство следует, очевидно, из того, что 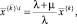 поскольку при
поскольку при 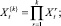
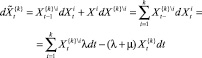
и, следовательно,
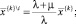
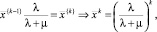 (8)
(8)
что приводит к нарушениям экспрессии генов за время T > 0, пропорциональное величине
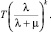 (9)
(9)
Таким образом, оптимизация работы генома предполагает за некоторое время T минимизацию
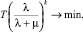 (10)
(10)
Однако полиплоидия (и это особенно наглядно демонстрирует поведение гепатоцитов) приводит к росту угрозы канцерогенеза, который можно в первом приближении представить в виде вероятности
1 – e–ρTk (11)
с задачей
1 – e–ρTk → min, (12)
где ρ – коэффициент канцерогенной активации генов на любой из k аллельных хромосом при k-полиплоидии.
Таким образом, при формировании полиплоидных структур (наряду с иными задачами и причинами её возникновения) решается задача оптимизации, аддитивно формирующаяся из подзадач (10) и (12):
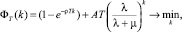 (13)
(13)
где A > 0 – некоторый, экспериментально (на основе эволюционного обоснования полиплоидии) определяемый коэффициент пропорциональности.
Заметим, что количество активных транспозонов существенно ниже их общего числа. Это приводит к соотношению λ << μ, а также к тому, что δ = –lnβ >> 1, поскольку общая генетическая масса активных транспозонов существенно меньше общего объема хромосомы.
Следовательно, задача (13) редуцируется в приближенную задачу
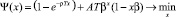 , (14)
, (14)
поскольку Ψ(x) = Φ(x) + ο(x).
Теорема. Решение задачи (14) существует и определяется трансцендентным уравнением
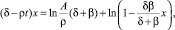
где δ = –lnβ.
Замечание. Решение не только существует, положительно, но и допускает оценку 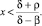
Рецензенты:
Андреев А.С., д.ф.-м.н., профессор, заведующий кафедрой информационной безопасности и теории управления, декан факультета математики и информационных технологий, Ульяновский государственный университет, г. Ульяновск;
Мищенко С.П., д.ф.-м.н., профессор, заведующий кафедрой алгебро-геометрических вычислений, Ульяновский государственный университет, г. Ульяновск.
Библиографическая ссылка
Бутов А.А., Карев М.А., Коваленко А.А., Кононова Г.В. АНАЛИЗ НАРУШЕНИЙ МЕТАБОЛИЗМА КАК СЛЕДСТВИЯ АКТИВИЗАЦИИ ТРАНСПОЗОНОВ В ПОЛИПЛОИДНЫХ КЛЕТКАХ // Фундаментальные исследования. 2015. № 2-27. С. 6030-6031;URL: https://fundamental-research.ru/ru/article/view?id=38613 (дата обращения: 19.06.2026).