



В настоящее время во всём мире актуальной представляется проблема сохранения редких и исчезающих локальных пород сельскохозяйственных животных [13, 22, 27, 28], адаптированных к экологическим условиям различных регионов [19, 30, 33, 36, 40]. Это сопряжено с широким распространением трансконтинентальных пород и приводит к увеличению генетического однообразия [21, 23]. Данная тенденция может привести к снижению резистентности животных к различным заболеваниям и сказаться на эффективности дальнейшей селекционной работы [17, 18, 25, 29, 32, 35].
На территории Российской Федерации имеет место широкий спектр природно-климатических условий, которые на протяжении длительного периода времени приводили к формированию уникальных сочетаний признаков локальных субпопуляций [26]. В этой связи закономерным видится появление генетического разнообразия, необходимого для совершенствования существующих, создания новых пород животных, оценки роли паратипических факторов в этом аспекте и процессе адаптации [7, 16, 20, 39]. Оценку генофонда чаще всего проводят по однонуклеотидному полиморфизму [34, 37, 41], генетическим системам сывороточных белков крови [3, 6, 7], частотам эритроцитарных антигенов [2, 4, 10, 42] и другим параметрам интерьера [15, 38].
Таким образом, целью наших исследований явилась сравнительная характеристика генофонда пород местного и интродуцированного скота Западной Сибири по частотам эритроцитарных антигенов.
Проведённые исследования являются частью программы комплексного изучения генофонда и фенофонда пород сельскохозяйственных животных Сибири [8, 11, 14, 24].
Материалы и методы исследований
Якутский и серый украинский скот был завезен ИЦиГ СО РАН в Западную Сибирь с целью сохранения генофонда пород [9, 12, 31]. У особей определяли частоты эритроцитарных антигенов с применением 46 моноспецифических сывороток по антигенам 8-ми генетическим системам.
Генетическое сходство оценивали с использованием коэффициентов Серебровского, Эвклида и кластерного анализа. В качестве метода группировки применяли алгоритм Уорда. Для поиска кластеров эритроцитарных антигенов, по частотам которых наблюдалось большее сходство, исходные данные подвергались транспонированию. Статистическую обработку проводили с помощью языка статистического программирования «R».
Результаты исследований и их обсуждение
Оценивался уровень генетического сходства интродуцированного и местного скота с красной степной породой, популяциями белого сибирского и якутского аборигенного скота [1, 27]. Характерной особенностью якутского и серого украинского скота явилось отсутствие у них антигенов K, I, P2, T2, B′, D′, J2′, O′, J′, B″, R1, X1, L′ и M.
Выявлен в целом высокий уровень генетического сходства всех сравниваемых пород животных. Наибольшее подобие установлено между интродуцированными породами (0,872 ± 0,051) по сравнению с остальными (0,711 ± 0,076 – 0,870 ± 0,053).
Использование многомерного агломеративного кластерного анализа позволило разбить субпопуляции крупного рогатого скота на группы и подгруппы по степени генетического сходства (рис. 1).
Полученные результаты показали наличие кластера 1, в котором оказались интродуцированные в Сибирь якутский скот и серая украинская порода. По всей видимости разведение завезённого на юг Западной Сибири якутского скота привело к изменению частот эритроцитарных антигенов по сравнению с аборигенным якутским. Предполагается, что подобные процессы имели место и в группах серого украинского скота и объясняются действием искусственного отбора, что встречается достаточно часто [1].
Наполнение кластера 2, с другой стороны, свидетельствовало о большем сходстве остальных пород. По всей видимости, это связано с тем, что чёрно-пёстрый, красный степной и белый сибирский скот на протяжении длительного периода времени разводились в условиях Сибири и были генетически более схожи между собой [5].
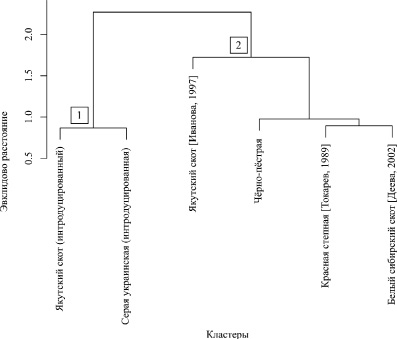
Рис. 1. Генетическое сходство пород крупного рогатого скота
Дальнейшее исследование частотного антигенного профиля проводилось по транспонированным исходным данным. Выявлен высокий уровень варьирования коэффициентов генетического сходства Серебровского (от 0,060 до 1). Количество коэффициентов выше 0,500 было более чем в 3,5 выше остальных, доля которых составляла 21,8 %. Это может служить показателем значительного сходства изучаемых субпопуляций и в большей мере по антигенам B2, O1, O2, E′1, E′2, G′, Q′, G″’, R2, W, V, L и Z. Отличия имелись лишь по антигенам H1 и F.
Используя имеющиеся транспонированные данные, прибегли к оценке генетического сходства с помощью дендрограммы и дистанции Эвклида (рис. 2).
Применение агломеративного метода кластеризации Уорда привело к появлению двух крупных кластеров. Состав первого кластера определялся антигенами с низкими частотами встречаемости, в то время как второй включал все остальные. Как можно судить по рисунку, каждый крупный кластер имел достаточно сложное деление на подгруппы. Так, в третьем подкластере можно обнаружить эритроцитарные антигены, частоты которых обладали большей изменчивостью по сравнению с таковыми в подкластере 4. В этих группах следует выделить антигены X1 и L′, по которым наблюдались самые высокие лимиты изменчивости (0,000–0,503 и 0,000–705). Значительное сходство обнаружено по антигенам B″’, R1, T2 и B′ с самой низкой концентрацией в подкластере 4.
Установлено высокое подобие по наиболее часто встречаемым антигенам подкластера 5. Среди представителей этой группы антигены F и H1 выделялись в большей степени (0,884–1,000 и 0,724–0,893 соответственно). В сравнении с ними антигены G2 и G3 отличались большим размахом вариации подкластера 6. По частотам других антигенов формирование кластеров поддавалось интерпретации в меньшей степени.
Примечательным представляется высокое подобие по частотам эритроцитарных антигенов скота, разводимого длительное время в условиях Сибири (чёрно-пестрый, красный степной и белый сибирский). Полученные результаты позволяют предполагать, что определённая степень сходства по изучаемым показателям у особей разных популяций одного вида может возникнуть при попадании их в одинаковые условия окружающей среды.
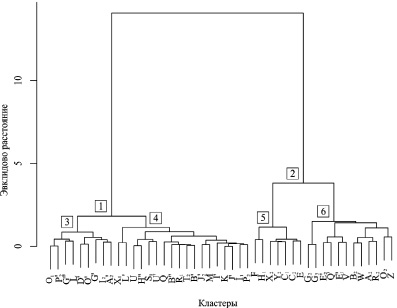
Рис. 2. Генетическое сходство пород крупного рогатого скота по частотам эритроцитарных антигенов
Заключение
Включение в соответствующую макрогруппу кластера 1 антигенов X1 и L′ на дендрограмме, по всей видимости, связано с их высокой встречаемостью у якутского аборигенного скота.
Предполагается, что в отношении частот антигенов G2, G3, X1 и L′ наблюдалось адаптивное преимущество. Делается предположение, что по прошествии длительного периода времени наблюдаемая кластерная структура исследуемых популяций скота может претерпеть значительные изменения. В отношении частот других антигенов выводы подобного рода формулировать не приходилось. Напротив, степень согласования частот эритроцитарных антигенов B″’, R1, T2 и B′, с одной стороны, а также F и H1 – с другой, была самой высокой. Это давало основание воспринимать частоты данных групп антигенов в качестве константных для всех сравниваемых пород скота.
Таким образом, выявлены комплексы антигенов, по которым имелось наибольшее сходство между разводимыми длительное время в условиях Сибири и интродуцированными и локальными субпопуляциями.
Рецензенты:
Клименок И.И., д.с.-х.н., профессор, зам. директора, ГНУ «Сибирский научно-исследовательский и проектно-технологический институт животноводства», п. Краснообск-1;
Дементьев В.Н., д.с.-х.н., профессор кафедры ветеринарной генетики и биотехнологии, ФГБОУ ВПО «Новосибирский государственный аграрный университет», г. Новосибирск.
Библиографическая ссылка
Камалдинов Е.В., Себежко О.И., Короткевич О.С. ФОНД ЭРИТРОЦИТАРНЫХ АНТИГЕНОВ КРУПНОГО РОГАТОГО СКОТА СИБИРИ // Фундаментальные исследования. 2015. № 2-19. С. 4197-4202;URL: https://fundamental-research.ru/ru/article/view?id=37929 (дата обращения: 30.06.2026).