



Экспериментирование с растительными популяциями с целью анализа реакции особей на изменение плотности посева растений представляет большой интерес для эволюционной теории, поскольку служит весьма эффективным способом познания закономерностей внутривидовой конкуренции – главного компонента борьбы за существование [8]. Растения на повышение популяционной плотности реагируют двумя способами: снижением вероятности выживания и пластическим изменением развития; пластичность реакции на повышение плотности можно рассматривать как один из механизмов сохранения генотипов в популяции, тогда когда гибель в результате увеличения плотности посева служит механизмом, обеспечивающим довольно быструю элиминацию определённых генотипов [9]. Выражением целостности растительного организма является скорреллированность его структур. Известно, что разные группы признаков характеризуются не только определённым уровнем варьирования, но также и различной степенью согласованности друг с другом. Анализ изменения силы и структуры связей отдельных признаков и их сопоставление с различиями по средним значениям и по размаху варьирования позволяют выявить специфику «поведения» отдельных признаков и их групп. Сильно варьирующие признаки с высоким уровнем детерминированности являются индикаторами адаптивной изменчивости организма [6].
Erysimum cheiranthoides L. (желтушник левкойный) в природных сообществах не образует зарослей. Он относится к однолетним сорным видам: встречается в нарушенных луговых и степных сообществах, а также в посевах культурных растений. E. cheiranthoides перспективен для выращивания в культуре как лекарственное растение кардиотонического действия [2].
Ранее нами изучено влияние плотности посева на средние значения морфологических признаков [3] и структуру корреляций в фазе цветения E. cheiranthoides [1]. Установлено, что при увеличении плотности посева уменьшались длина флоральной части побега, число и общая длина боковых побегов (паракладиев), число плодов (стручков), число семян на особь, масса плодов, масса семян, масса побегов, масса корней и масса особи; длина префлоральной части побега увеличивалась, а число семян в стручке, длина главного побега и репродуктивное усилие (отношение массы плодов к общей фитомассе особи) не изменялись. Цель настоящего исследования – установить влияние плотности посева на изменчивость корреляционных связей изученных морфологических признаков E. cheiranthoides в фазе плодоношения.
Материал и методы исследования
Опытные посевы проводили семенами пятой репродукции ранней весной. Семена высевали рядовым способом, с междурядьями 70 см на ровных участках площадью 60 погонных метров для каждого варианта. Семена высевали на одинаковую глубину, точно по счету, на каждый погонный метр. Испытывали 5 вариантов: 1 – 3000 шт. (масса 0,66 г) очень высокой; 2 – 1000 шт. (0,22 г) высокой; 3 – 500 шт. (0,11 г) средней, 4 – 200 шт. (0,044 г) низкой плотности посева и 5 – 50 шт. (0,011 г) – разреженный посев. Сорняки в течение сезона уничтожали. В каждом варианте посева отбирали в фазе плодоношения (молочно-восковая спелость семян) по 50 особей смешанным способом: с 5 участков по 10 погонных метров случайным способом отбирали по 10 растений. Для анализа корреляционных связей использовали метод корреляционных плеяд [7]. Корреляционные кольца построены на уровне связей r > 0,7. Сравнение матриц корреляций проводили по методу, предложенному Ростовой [6]. При анализе корреляционных связей использовали следующую шкалу уровней связи: слабая связь r < 0,3, умеренная 0,3 < r < 0,5, значительная 0,5 < r < 0,7, сильная 0,7 < r < 0,9, очень сильная r > 0,9.
Результаты исследования и их обсуждение
Из числа изученных признаков наиболее изменчивы (CV=49–186 %) и высоко детерминированы (R2ch=0,293–0,659) следующие: число и общая длина паракладиев (В, Г), число стручков (Д), общее число семян (И), масса побегов (К), масса стручков (Л), масса корней (М), масса семян (Н), масса особи (О) (таблица). У всех этих признаков, характеризующих продуктивность особей, при увеличении плотности посева происходило снижение их средних значений [3]. Для многих однолетних видов характерно увеличение вариабельности размеров при увеличении плотности посева [9]. Нами также отмечена высокая степень дифференциации особей по размерам у E. cheiranthoides в посевах высокой плотности [2]. Мощные особи по общей длине боковых побегов (паракладиев) превышали слабо развитые растения в 500 раз, по массе особи и массе семян – в 32 раза. Такая высокая вариабельность признаков свидетельствует о больших приспособительных возможностях вида. Благодаря этой особенности данный вид может расселяться как сорное растение в посевах пропашных культур [5].
Общая и согласованная изменчивость признаков в фазе плодоношения у Erysimum cheiranthoides в посевах разной плотности
|
Признаки |
Плотность посева |
FD1 |
|||||||||
|
1 |
2 |
3 |
4 |
5 |
|||||||
|
CV |
R2ch |
CV |
R2ch |
CV |
R2ch |
CV |
R2ch |
CV |
R2ch |
||
|
Длина префлоральной части побега, см |
56.2 |
0,231 |
54,1 |
0,263 |
82,2 |
0,279 |
114,3 |
0,287 |
134,3 |
0,355 |
91,8 |
|
Длина флоральной части побега, см |
48,8 |
0,331 |
55,1 |
0,434 |
37,5 |
0,257 |
33,0 |
0,272 |
22,1 |
0,143 |
78,2 |
|
Число паракладиев, шт. |
136,3 |
0,448 |
176,0 |
0,604 |
131,4 |
0,588 |
106,3 |
0,580 |
61,4 |
0,491 |
93,7 |
|
Общая длина паракладиев, см |
159,0 |
0,487 |
186,3 |
0,613 |
137,4 |
0,589 |
117,3 |
0,586 |
63,8 |
0,451 |
94,9 |
|
Число стручков, шт. |
111,1 |
0,542 |
165,0 |
0,616 |
110,4 |
0,609 |
102,6 |
0,601 |
55,7 |
0,539 |
92,2 |
|
Число семян в стручке, шт. |
16,8 |
0,096 |
17,5 |
0,165 |
15,0 |
0,055 |
13,7 |
0,063 |
8,0 |
0,032 |
71,1 |
|
Общее число семян, шт. |
116,2 |
0,508 |
167,9 |
0,609 |
121,2 |
0,584 |
102,2 |
0,622 |
55,0 |
0,535 |
81,1 |
|
Масса побегов, г |
114,0 |
0,502 |
125,4 |
0,651 |
99,5 |
0,619 |
106,2 |
0,602 |
55,8 |
0,403 |
95,0 |
|
Масса стручков, г |
101,1 |
0,514 |
122,9 |
0,648 |
95,5 |
0,588 |
94,7 |
0,607 |
51,0 |
0,498 |
90,9 |
|
Масса корней, г |
149,7 |
0,329 |
121,7 |
0,591 |
98,8 |
0,520 |
100,8 |
0,551 |
68,9 |
0,293 |
96,6 |
|
Масса семян, г |
131,7 |
0,340 |
168,3 |
0,609 |
122,4 |
0,584 |
101,8 |
0,623 |
55,0 |
0,535 |
85,9 |
|
Масса особи, г |
127,1 |
0,415 |
122,8 |
0,659 |
96,5 |
0,613 |
97,8 |
0,630 |
48,9 |
0,516 |
95,5 |
|
Репродуктивное усилие |
21,4 |
0,101 |
15,1 |
0,035 |
16,2 |
0,045 |
13,7 |
0,101 |
19,4 |
0,132 |
58,7 |
|
Длина главного побега, см |
28,2 |
0,110 |
26,5 |
0,164 |
20,1 |
0,057 |
24,6 |
0,098 |
21,6 |
0,170 |
81,8 |
|
Итого |
94,1 |
0,354 |
108,9 |
0,476 |
84,6 |
0,427 |
80,7 |
0,445 |
51,5 |
0,364 |
- |
Условные обозначения: CV – коэффициент вариации ( %), R2ch – коэффициент детерминации, FD1 – степень сходства структуры связей признаков ( %).
Снижение коэффициентов детерминации и вариации вышеперечисленных признаков продуктивности происходит в 1-м и 5-м вариантах плотности посевов. В условиях 2–4 вариантов посева уровень детерминированности и вариабельности этих признаков был выше. Для большинства этих признаков в посевах разной плотности характерна высокая стабильность структуры связей (FD1=91–97 %) (таблица). Исключение составили два признака – число и масса семян (FD1=81–86 %), что связано со снижением уровня связей этих признаков с массой особи в 1-м варианте плотности посева (рисунок).
В условиях очень высокой плотности посева (вариант 1) ослабевали связи у большинства признаков продуктивности по сравнению с вариантами 2–4. Более существенное ослабление корреляционных связей (0,7 < r < 0,8) наблюдалось между признаками побега (В, Г, И, К) и признаками семенной продуктивности (Д, Л, И) с массой особи (О) (рисунок). В варианте 5 корреляционные связи признаков побега и признаков семенной продуктивности с массой особи усиливались по сравнению с вариантом 1. Вариант 5 отличался от вариантов 2-4 ослаблением корреляционных связей признаков побега (В, Г) с массой особи и массы побегов (К) с числом побегов (Г). 1-й и 5-й варианты посева отличались от вариантов 2–4 более слабой связью массы корней (М) с признаками продуктивности. Если в вариантах 2-4 связь массы корней с признаками продуктивности была очень сильной (r > 0,9) и сильной (r > 0,8), то в вариантах 1 и 5 она снижалась до значительной (0,5 < r < 0,7) и умеренной (0,3 < r < 0,5).
Длина префлоральной (А), флоральной части (Б) и общая длина главного побега (Р) по-разному реагируют на изменения плотности посевов [3]. Чем меньше средние значения длины префлоральной части побега, тем выше средние значения признаков продуктивности. Детерминированность и вариабельность длины префлоральной части побега уменьшается по мере увеличения плотности посева. При снижении плотности посева от 1-го варианта к 5-му размах варьирования признаков продуктивности, как показано выше, снижается, а вариабельность длины префлоральной части побега возрастает (таблица). Связь с признаками продуктивности в разных вариантах значительная (–0,5 > r > –0,7), а в разреженном посеве – сильная (r < –0,7). Структура связей длины префлоральной части побега в посевах разной плотности стабильна (FD1=91,8 %).
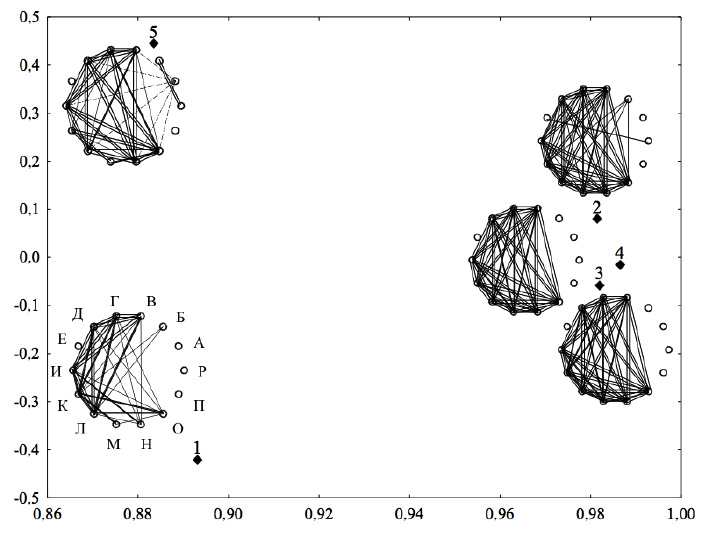
Сравнение матриц корреляций морфологических признаков E. cheiranthoides в посевах разной плотности. По оси Х – фактор сходства матриц, по оси Y – фактор специфичности матриц. Сечения цилиндров – на уровне связи r > 0,7. Обозначения корреляций: сплошные линии – положительные, пунктирные – отрицательные корреляции; 0,7 < r < 0,8 – тонкая линия, 0,8 < r < 0,9 – жирная линия, r > 0,9 – двойная жирная линия. Названия признаков даны в методике
При увеличении плотности посева длина флоральной части побега (Б) уменьшается, детерминированность и вариабельность этого признака увеличиваются. Корреляционные связи длины флоральной части побега с признаками продуктивности значительные (0,5 < r < 0,7), кроме разреженного посева. В разреженном посеве корреляции с признаками продуктивности сильно ослабевают (–0,2 < r < 0,3), а связь с длиной главного побега, наоборот, усиливается и становится очень сильной (r > 0,9). В разреженном посеве длина флоральной части побега составляет 97,2 % от длины главного побега. Структура связей длины флоральной части побега в посевах разной плотности менее стабильна, чем у признаков продуктивности (FD1=78,2 %).
Слабо выражена реакция на изменения плотности посева у длины главного побега (Р). Уровень детерминированности и вариабельности этого признака в посевах разной плотности остаются низкими. В разреженном посеве его детерминированность увеличивается за счет усиления связи (r > 0,9) с длиной флоральной части побега (рисунок). Структура связей менее стабильна (FD1=82 %), чем у признаков продуктивности.
К признакам, слабо детерминированным (R2ch=0,032–0,165) и слабо изменчивым (CV=8,0-21,4 %), относятся число семян в стручке (Е) и репродуктивное усилие (П). Число семян в стручке при увеличении плотности посева уменьшается незначительно, а уровень детерминированности и вариабельности повышаются также незначительно. Структура связей менее стабильна (FD1=71 %), чем у признаков продуктивности. Корреляции с признаками продуктивности во всех вариантах плотности очень слабые (r < 0,1). В загущенных посевах усиливается связь числа семян в стручке с длиной главного побега (Р). Она становится значительной (0,5 < r < 0,7) в первом варианте и сильной (r > 0,8) во втором варианте плотности посева.
Репродуктивное усилие под влиянием плотности посева изменяется незначительно. Известно, что у однолетних видов этот показатель стабилен [4]. Детерминированность и вариабельность признака низкие, связи с признаками продуктивности слабые (r > –0,3). В 1-м и 5-м вариантах плотности посева происходит усиление связей репродуктивного усилия с длиной флоральной части побега и длиной главного побега (0,5 < r < 0,7). В остальных вариантах эта связь слабая (r < 0,3). Структура связей в посевах разной плотности нестабильная (FD1=59 %).
Матрицы вариантов 1 и 5 характеризуются более низким уровнем связей, чем матрицы вариантов 2-4 (рисунок). Кроме того, средний коэффициент детерминации по всей матрице (R2m) в вариантах 1 и 5 примерно одинаковый, что связано с тем, что в варианте 5 число очень высоких (r > 0,9) и слабых (–0,3 < r < 0,3) связей больше, чем варианте 1.
Сходство структуры связей по полным матрицам высокое и составляет 89,6 %. Вычисление коэффициентов корреляции между матрицами (rz) показало, что наиболее сходны по структуре связей матрицы 2-4 (rz=0,956–0,968). Значительные различия обнаруживаются между 1 и 5 вариантами (rz=0,624). 1-й вариант от 2–4 (rz=0,832–0,879) и 5-й от 2–4 (rz=0,824–0,887) отличаются менее существенно. Ослабление корреляционных связей в крайних вариантах плотности между признаками продуктивности можно объяснить разными темпами онтогенеза особей. Многолетние наблюдения показали, что в условиях очень высокой плотности посева растения E. cheiranthoides переходят в генеративную фазу раньше, чем в разреженных посевах. Такую закономерность мы объясняем тем, что степень ветвления побега в разреженном посеве у растений увеличивается более, чем в 10 раз. Число боковых побегов увеличивается в 13,5 раза. Каждый боковой побег сначала развивает вегетативную (префлоральную часть) и значительно позднее генеративную (флоральную часть). В совокупности боковые побеги формируют флоральную часть главного побега, число метамеров до цветения у растений в условиях разреженного посева значительно больше, чем у особей в условиях высокой плотности посева. В условиях очень высокой плотности посева у растений формируется простое соцветие кисть или сложное – двойная кисть, в условиях разреженного посева – тройная кисть или кисть более высокого порядка.
Заключение
В условиях культуры изучено влияние плотности посева на изменчивость корреляционных связей морфологических признаков у Erysimum cheiranthoides L. Испытывали 5 вариантов плотности посева: очень высокой (1 вариант), высокой (2 вариант), средней (3 вариант), низкой (4 вариант) плотности и разреженный посев (5 вариант). Корреляционные связи стабильных признаков – число семян в стручке, репродуктивное усилие изменялись незначительно. Пластичные признаки – число и общая длина боковых побегов, число плодов, число семян на особь, масса побегов, масса плодов, масса семян масса корней, масса особи оставались высоко детерминированными и сильно варьирующими. Разнонаправленный характер изменчивости корреляционных связей обнаруживался у признаков побега – длина префлоральной и флоральной части и общая длина. При увеличении плотности посева детерминированность и вариабельность длины префлоральной части побега уменьшались, длины флоральной части побега увеличивалась, длины всего побега оставались относительно стабильными. По структуре связей наиболее сходны корреляционные матрицы 2-4 вариантов. Наибольшие различия по структуре связей отмечены между матрицами 1 и 5. Матрицы вариантов 1 и 5 отличаются более низкими уровнями связей от матриц вариантов 2–4.
Рецензенты:
Викторов В.П., д.б.н., профессор, кафедра ботаники ФГБОУ ВПО «МПГУ», г. Москва;
Марков М.В., д.б.н., профессор, кафедра физической географии и экологии ФГБОУ ВПО «ТвГУ», г. Тверь.
Работа поступила в редакцию 29.07.2014.
Библиографическая ссылка
Годин В.Н., Гонтарь Э.М. ВЛИЯНИЕ ПЛОТНОСТИ ПОСЕВА НА КОРРЕЛЯЦИИ ПРИЗНАКОВ В ФАЗЕ ПЛОДОНОШЕНИЯ У ERYSIMUM CHEIRANTHOIDES L // Фундаментальные исследования. 2014. № 8-7. С. 1586-1590;URL: https://fundamental-research.ru/ru/article/view?id=35257 (дата обращения: 04.08.2026).