



При интродукции хвойных в новые для них экологические условия надежным диагностическим признаком состояния древесных растений служит качественный и количественный состав пигментной системы – хлорофиллов и каротиноидов – главных фоторецепторов фотосинтезирующей клетки. При успешной интродукции содержание хлорофилла интродуцента очень близко к содержанию его у местной породы.
Природные условия юга Приморского края определяются муссонным климатом. Это обусловливает весьма специфичные климатические условия, которые существенно влияют на успешность интродукции. Зимой массы холодного и сухого воздуха перемещаются с континента к морю, а летом наблюдается обратный процесс [1]. В таких условиях затяжная весна, пасмурное, с большим количеством туманов и осадков начало лета, суровая (сравнительно с европейской частью страны) с холодными северными ветрами зима оказывают лимитирующее действие на нормальное развитие растений-интродуцентов. Кроме того, гористый рельеф Приморья обусловливает формирование различных микроклиматических условий даже в пределах небольшой территории. Высота над уровнем моря, экспозиция склона, относительная высота над дном долины, форма склона и долины оказывают существенное влияние на перераспределение тепла и влаги.
В таких условиях выживание интродуцентов возможно только при их успешной адаптации к комплексу природно-климатических условий района интродукции [2].
Целью настоящего исследования явилась сезонная количественная оценка параметров накопления фотосинтетических пигментов в хвое четырех интродуцированных видов рода Ель и выявление на биохимическом уровне адаптивных механизмов интродуцента в условиях дендрария Горнотаежной станции ДВО РАН.
Материалы и методы исследования
Объектом исследования явились хвойные породы в возрасте 45–50 лет дендрария Горнотаёжной станции ДВО РАН (Уссурийский район, Приморский край). Изучалась годовая динамика накопления фотосинтетических пигментов в хвое второго года у интродуцированных видов ели колючей (Picea pungens Engelm.), ели Энгельмана (Picea engelmanni (Parry) Engelm.), ели шероховатой (Picea asperata Mast.), ели Мейера (Picea meyeri Rehd. ex Wils.). Для сравнения в качестве эталона исследовали динамику накопления пигментов у дальневосточного вида – ели корейской (Picea koraiensis Nakai).
Ель корейская – дерево до 30 м высотой с пирамидальной кроной и поникающими ветвями. Естественно произрастает на Дальнем Востоке, распространена по всему Приморскому краю и в южных районах Хабаровского края, южнее 49°с.ш. Произрастает обычно на щебнистых увлажненных почвах по долинам рек, вместе с елью аянской и пихтой белокорой. Хвоинки 9–22 мм длиной и 1,5–1,8 (–2,2) мм шириной, четырехгранные, длинно заостренные (с острием 0,5–0,7 мм), зеленые, с 2–4 устьичными линиями на каждой из сторон. Обладает высокой устойчивостью к неблагоприятным факторам среды.
Eль колючая – дерево до 30–45 м высотой, со стволом 70–120 см в диаметре, с симметричной конусовидной кроной из горизонтально мутовчатых ветвей, доходящих при изолированном стоянии до земли. Хвоинки 20–30 мм, 1–1,5 мм шириной, четырехгранные, плотные и сильно колючие, от зеленых до серебристо-беловатых, с 3–5 белыми устьичными линиями на каждой стороне, с беловатым налетом или без него, отстоящие от побега; хвоя держится 4–6 (–9) лет. В природе встречается одиночно или небольшими группами вдоль рек, по северным склонам гор западных районов Северной Америки. Растет в горах на высоте 2000–3300 м над уровнем моря. Декоративна в любое время года. Среди многочисленных представителей рода выделяется стройностью и красотой, нетребовательностью к условиям произрастания, морозостойкостью и устойчивостью к воздушным загрязнениям, превосходя по этому показателю многих собратьев.
Ель Энгельмана – дерево до 30–50 м высотой, со стволом до 90 см в диаметре, с густой конусовидной кроной (может быть узкой и острой) и слегка поникающими ветвями. Хвоинки 15–25 (–30) мм, 1,5–2 мм шириной, четырехгранные, острые, достаточно жесткие (но мягче, чем у ели колючей), прямые или слегка изогнутые, сизо-зеленые, направленные к верхушке побега; хвоя держится 5–10 (–15) лет. Зимостойка и вынослива в отношении неблагоприятных климатических факторов. Неприхотлива к почве, считается медленно растущей. Родиной является Запад Северной Америки, в чистых и смешанных лесах, на высоте 1500–3500 м над уровнем моря.
Ель шероховатая – малоизвестный в нашей стране вид из западных районов Китая, где произрастает в зоне субальпийских лесов, на высотах 2700–3500 м. Дерево до 25–45 м высотой, с конической кроной. Хвоинки 10–15 (–20) мм длиной, 1–1,8 мм шириной, четырехгранные, с 4–8 устьичными линиями на каждой из сторон. Засухоустойчива, хорошо развивается в условиях города. По декоративности не уступает ели колючей и ее формам.
Ель Мейера – дерево до 30 м высотой с ширококолонновидной или конической негустой кроной, близкородственный вид ели шероховатой. От неё отличается несколько менее крупными, более симметричными шишками с закругленными по верхнему краю чешуями, а также туповатыми на верхушке хвоинками (у P. asperata – острые) и почками, которые имеют одну окраску с побегами. Распространена в Северном Китае [5].
Количество хлорофиллов а, b и сумму каротиноидов определяли спектрофотометрическим методом с использованием методических разработок [6]. Образцы хвои второго года вегетации с постоянной навеской 0,2 г отбирали в трехкратной повторности в течение года. Далее в лабораторных условиях получали вытяжку пигментов в ацетоне. Экстракты пигментов фильтровались вакуумным способом. Оптические плотности пигментных вытяжек определяли с помощью однолучевого автоматизированного спектрофотометра СФ-56 (ЛОМО) по центрам поглощения: для хлорофиллов а и b – 644 и 662 нм, для каротиноидов – 440,5 нм.
Основой расчета концентрации пигментов хлоропластов служили формулы Веттштейна для 100 %-го ацетона:
Са = 9,78∙D662 – 0,99∙D644;
Сb = 21,42∙D644 – 4,65∙D662;
Са+b = 5,13∙D662 + 20,43∙D644;
Скар = 4,69∙D440,5 – 0,268∙(Са+–b),
где С – концентрация хлорофиллов а, b и каротиноидов в мг/л; D – оптическая плотность в центрах поглощения пигментов 440,5; 644 и 662 нм.
Содержание пигментов в хвое вычислялось по формуле: А = (С∙V)/(P∙1000), где А – содержание пигмента в мг на 1 г сырой навески; С – концентрация пигмента в мг/л (после расчета по формулам (1), (2), (3), (4); V – объем вытяжки пигмента (мл); Р – навеска хвои (г).
Результаты исследования и их обсуждение
Содержание пигментов в течение периода вегетации является довольно динамичным показателем, что отмечалось многими авторами [3].
Результаты проведенных исследований позволили установить, что содержание суммы зеленых и желтых пигментов в хвое интродуцентов подвержено значительным сезонным колебаниям: ель колючая – от 1,24 до 2,20 мг/г сырого веса, ель Энгельмана – от 1,02 до 1,69 мг/г, ель шероховатая – от 0,94 до 1,90 мг/г, ель Мейера – от 0,98 до 2,33 мг/г.
Установлено, что фонд пигментов у ели колючей активно пополнялся в летний период с мая по июнь с максимальными значениями 2,20 мг/г сырого веса и в сентябре – октябре (рис. 1, а). Сравнительный анализ уровня пигментов показал, что по суммарному содержанию ель колючая не только не уступала местной ели корейской, но и опережала накопление пигментов в 1,10 раз.
Близость количественных характеристик и сезонные закономерности накопления пигментов для трех интродуцированных елей хорошо прослеживаются на рис. 1, б,в,г.
Согласно полученным результатам, для ели Энгельмана, ели шероховатой и ели Мейера максимальные значения накопления хлорофиллов и каротиноидов приходятся на осенний период – октябрь – ноябрь, в то время как у дальневосточной ели корейской наибольшее содержание пластидных пигментов отмечено в январе – феврале.
Разница в суммарном количестве пигментов у интродуцента в сравнении с местным видом определяется защитными функциями пигментов. Как правило, максимальное их содержание наблюдается в наиболее критические погодно-климатические периоды, что подтверждает их защитные функции [4].
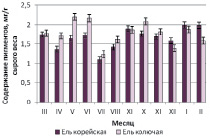
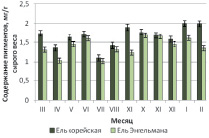
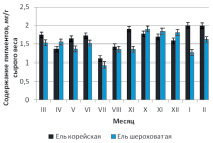
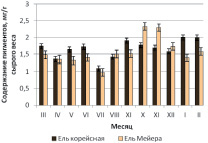
Рис. 1. Сезонная динамика суммарного содержания фотосинтетических пигментов (хлорофиллы а, b и каротиноидов) хвои в паре «местный вид – интродуцент»: а – ель корейская – ель колючая; б – ель корейская – ель Энгельмана; в – ель корейская – ель шероховатая; г – ель корейская – ель Мейера
Исследование процессов сезонного накопления желтых пигментов – каротиноидов ‒ выявило следующие периоды накопления фонда: для дальневосточного вида они приходятся на осенне-зимний период времени года, с максимумом в феврале – 0,33 мг/г.
Проанализировав содержание каротиноидов у интродуцированных елей установили, что ель колючая активно синтезирует желтые пигменты весной (март – апрель) и в конце осени (октябрь – ноябрь); ель Энгельмана и ель Мейера имеют осенние максимальные значения (0,28 и 0,33 мг/г соответственно), а для ели шероховатой характерно равномерное накопление пигмента осенью (октябрь – 0,30 мг/г) и в начале зимы (декабрь – 0,29 мг/г). Концентрация каротиноидов в начале лета незначительная у всех видов, минимум ее наблюдается в период цветения обоих видов елей, в конце мая – начале июня (рис. 2).
В отличие от зеленых пигментов количество каротиноидов в осенне-зимний период имеет тенденцию к росту. Высокие показатели значений каротиноидов осенью и зимой обусловлены повышенным уровнем инсоляции, наиболее характерным для климатических условий Приморского края. В этот период желтые пигменты в большей степени выполняют защитную функцию – сохраняют хлорофиллы от избытка солнечной радиации, в остальное же время они действуют как пигменты, дополнительно поглощающие свет для фотосинтетических процессов.
Среднегодовое количество пластидных пигментов в двухлетней хвое ели корейской, произрастающей в естественных для нее условиях и 4-х интродуцированных в дендрарий видов приведено в таблица.
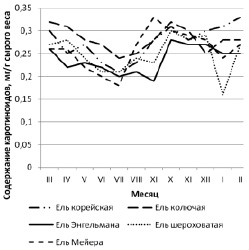
Рис. 2. Сезонная динамика содержания каротиноидов в хвое видов Picea
Среднегодовое количество хлорофиллов а, b и каротиноидов и их соотношение в хвое Picea
|
Название вида |
Содержание пигментов, мг/г сырой навески |
Отношение |
||||
|
Аа |
Аb |
Акар |
Аа+b+кар |
а/b |
Sа+b/Sкар |
|
|
Ель корейская |
0,92 ± 0,03 |
0,46 ± 0,02 |
0,27 ± 0,02 |
1,65 ± 0,04 |
2,00 |
5,11 |
|
Ель колючая |
0,99 ± 0,03 |
0,51 ± 0,04 |
0,28 ± 0,01 |
1,78 ± 0,05 |
1,94 |
5,36 |
|
Ель Энгельмана |
0,73 ± 0,02 |
0,41 ± 0,05 |
0,24 ± 0,02 |
1,41 ± 0,08 |
1,85 |
4,88 |
|
Ель шероховатая |
0,83 ± 0,04 |
0,43 ± 0,03 |
0,25 ± 0,04 |
1,51 ± 0,11 |
1,93 |
5,04 |
|
Ель Мейера |
0,86 ± 0,02 |
0,46 ± 0,04 |
0,26 ± 0,03 |
1,58 ± 0,07 |
1,87 |
5,08 |
Согласно полученным данным, инорайонные виды елей по основным показателям пигментного комплекса не только не уступают, но и превосходят местные виды. Так, содержание хлорофиллов и каротиноидов в хвое ели колючей (1,78 мг/г) выше, чем у дальневосточной ели корейской (1,65 мг/г). Общий фонд пигментов у ели Мейера (1,58 мг/г) приближен к сумме зеленых и желтых пигментов у местного вида (1,65 мг/г), что может свидетельствовать об успешной адаптации интродуцентов к условиям дендрария Горнотаежной станции.
Одним из информативных показателей, характеризующих работу фотосинтетического аппарата, является отношение хлорофилла а к хлорофиллу b. Это отношение связано с активностью «главного» хлорофилла а: чем оно больше, тем интенсивнее фотосинтез. В норме этот показатель должен соответствовать 2,0–3,0. Стабильность в работе фотосинтетического аппарата мы наблюдаем у местного вида (2,00), тенденцию к нормальному функционированию фотосинтетического комплекса отмечена у ели колючей и ели шероховатой.
Отношение суммы хлорофиллов к каротиноидам (a + b/каротиноиды) играет не менее важную роль при характеристике работы фотосинтетического аппарата. Это соотношение в норме стабильно и очень чутко реагирует на изменения различных факторов среды. В целом приведенные данные (таблица) свидетельствуют о сходстве в характере формирования хлорофиллов и каротиноидов в хвое дальневосточного и интродуцируемого вида.
Выводы
Содержание хлорофиллов а, b и каротиноидов в хвое интродуцируемых видов подвержено сезонным изменениям, что объясняется их биологическими особенностями.
По уровню активности фотосинтетического аппарата интродуцированные виды располагаются в следующем порядке (по убыванию): ель колючая → ель Мейера → ель шероховатая → ель Энгельмана.
Близость количественных характеристик пигментного комплекса интродуцентов (ель колючая и ель Мейера) и дальневосточного вида (ель корейская) свидетельствует в пользу мнения об адаптационной сбалансированности фотосинтетического аппарата инорайонных видов к световому режиму, климатическим и природным условиям дендрария Горнотаёжной станции.
Полученные материалы могут быть использованы для прогнозирования успешности процесса интродукции различных видов Picea.
Список литературы
1. Агроклиматический справочник по Приморскому краю; под ред. В.В. Белевич. – Л.: ГИМИЗ, 1960. – 130 с.
2. Базилевская Н.А. Теории и методы интродукции растений – М.: Изд-во Московского ун-та, 1964. – 131 с.
3. Николаевский В.С. Экологическая оценка загрязнения среды и состояния наземных экосистем методами фитоиндикации. – М.: МГУЛ, 1998. – 130 с.
4. Петухова И.П. Эколого-физиологические основы интродукции древесных растений. – М.: Наука, 1981. – С. 76–78.
5. Фирсов Г.А., Орлова Л.В. Хвойные в Санкт-Петербурге. – СПб.: ООО Изд-во «Росток», 2008. – 336 с.
6. Шлык А.А. Определение хлорофиллов и каротиноидов в экстрактах зеленых листьев // Биохимические методы в физиологии растений. – М.: Наука, 1971. – С. 154–170.
References
1. Agroklimaticheskij spravochnik po Primorskomu kraju (Agroclimaticale directory of the Primorsky region. ed. V.V. Belevich. GIMIZ, 1960. 130 p.
2. Bazilevskaja N.A. Teorii i metody introdukcii rastenij (Theory and methods of plant introduction). Moscow University Press, 1964. 131 p.
3. Nikolaevskyi V.S. Jekologicheskaj ocenka zagrjaznenija sredy I sostojanija nazemnyh jekosistem metodomi fitoindikacii (Ecological estimation of water pollution and terrestrial ecosystems condition by phytoindication methods). Moscow, MGUL, 1998. 130 p.
4. Petukhova I.P. Jekologofiziologicheskie osnovy introdukciid revesnyh rastenij (Ecological and physiological basis of the introduction of woody plants ). Moscow: Nauka, 1981, pp. 76–78.
5. Firsov G.A., Orlova L.V. Hvojnye v Sankt-Peterburge (Conifers in St. Petersburg) Publishing «Rostok». SPb., 2008. 336 p.
6. Shlyk A.A. Opredelenie hlorofillov i karotinoidov v jekstraktah zelenyh listev (Determination of chlorophylls and carotenoids in green leaves extracts). Biochemical methods in plant physiology. Moscow, Nauka, 1971, pp. 154–170.
Рецензенты:
Зориков П.С., д.б.н., профессор, директор Горнотаежной станции им. В.Л. Комарова ДВО РАН, г. Уссурийск;
Гуков Г.В., д.с.-х.н., профессор, академик МААО, заслуженный работник высшей школы, Приморская государственная сельскохозяйственная академия, кафедра лесоводства, г. Уссурийск.
Работа поступила в редакцию 06.11.2013.
Библиографическая ссылка
Титова М.С. ОСОБЕННОСТИ ФОТОСИНТЕЗИРУЮЩЕЙ АКТИВНОСТИ ХВОИ ИНТРОДУЦИРОВАННЫХ ВИДОВ PICEA А. DIETR. В ДЕНДРАРИИ ГОРНОТАЕЖНОЙ СТАНЦИИ // Фундаментальные исследования. 2013. № 11-1. С. 128-132;URL: https://fundamental-research.ru/ru/article/view?id=33022 (дата обращения: 02.07.2026).