



Выбор рыбами мест обитания определяется целым рядом экзогенных и эндогенных факторов. В естественной среде довольно часто оптимальные для жизнедеятельности величины одного фактора могут не совпадать с таковыми по другим. Так, например, несмотря на обилие кормовых организмов, они могут быть слабо доступны для молоди рыб вследствие высокого пресса хищников (Fraser, Gilliam, 1987). Также возможны ситуации, когда в зонах с оптимальными для роста и развития рыб температурами кормовые организмы малочисленны или отсутствуют (Wurtsbaugh, Neverman, 1988). Таким образом, случай «идеального местообитания», то есть участка водоема, где существует совокупность оптимальных значений различных факторов окружающей среды, относительно редок. В связи с этим остается открытым вопрос, какие из факторов являются доминирующими при заселении рыбами тех или иных биотопов. Считается, что рыбы будут сосредотачиваться в тех зонах, в которых энергетические затраты на поддержание жизнедеятельности будут минимальны, а рост и развитие – максимальны. Как было показано ранее, в таких местообитаниях значения температур воды должны приближаться к видовому оптимуму (Jobling, 1981).
Важность укрытий для жизнедеятельности молоди различных видов рыб неоднократно исследовалась как в естественной, так и в экспериментальной среде (Christensen and Persson, 1993; Persson, 1993; Bevelhimer, 1996 и др.). При этом было показано, что присутствие или отсутствие укрытий может заметно влиять на поведение рыб и выбор ими мест для обитания. Однако за редким исключением данные исследования выполнялись без учета температурных требований рыб (Bevelhimer, 1996). В связи с этим целью данной работы было установление температурных предпочтений молоди речного окуня при наличии и отсутствии укрытий в экспериментальной среде.
Материалы и методы исследования
Экспериментальный материал, сеголетки (0+) окуня, был выловлен в прибрежье Рыбинского водохранилища в сентябре 2012 г. Далее молодь содержали в общем акклимационном аквариуме (температура 15 °С). Рыб кормили один раз в сутки личинками хирономид. Средняя длина молоди составляла 70,3 ± 1,4 мм, средняя масса – 6,6 ± 0,4 г.
Эксперименты были выполнены в двух лотках горизонтальной термоградиентной установки. Температурный градиент устанавливался от 15 до 30 °С. На начало опыта в отсеках первого лотка установки находились укрытия (группа I), во втором лотке укрытия отсутствовали (группа II). Рыбы помещались в отсек с температурой, равной температуре акклимации (15 °C). После достижения рыбами зоны окончательных избираемых температур (ОИТ) из первого лотка (группа I) изымали укрытия, расположенные в трех отсеках ближайших к зоне температурного оптимума (7,8 и 9 отсеки). Во второй лоток (группа II) добавляли укрытия в два отсека, расположенные по сторонам от зоны оптимума (6 и 10 отсеки). Спустя четверо суток, были повторно изъяты из первого лотка укрытия (6 и 10 отсеки), а во втором лотке они были передвинуты в 5 и 11 отсеки.
Наблюдения за рыбами производились в светлое время суток с использованием видеосъемки с последующей ее обработкой на компьютере. Распределения рыб в отсеках термоградиентной установки отмечалось 18 раз в течение светлого времени суток. Молодь кормили один раз в сутки (в 12.00), при этом корм помещался в отсеки, в которых находилась рыба на момент кормления.
На основе полученных данных были рассчитаны значения средней избираемой температуры за сутки. Значения ОИТ рассчитывались исходя из отсутствия достоверных различий в среднесуточных значениях избираемых температур в течении трех последовательных суток. Статистическое сравнение выборок проводилось с использованием критерия Стьюдента. Для статистических расчетов использовался программный пакет Statistica.
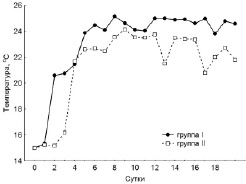
Рис. 1. Среднесуточные значения температур, предпочитаемых молодью окуня. Группа I ‒ в присутствии укрытий, группа II ‒ при отсутствии укрытий
Результаты исследования и их обсуждение
Полученные данные показывают, что наличие либо отсутствие в экспериментальной среде укрытий влияет на поведение рыб в температурном градиенте. Адаптация молоди II группы к условиям термоградиентной установки при отсутствии укрытий происходила гораздо медленнее. Рыбы дольше находились в отсеке, в который их поместили в начале эксперимента (рис. 1). Лишь на 4-е сутки значения температур предпочитаемых рыбами обеих групп выровнялись (21,5 группа I и 21,7 группа II). Далее молодь I группы продолжала выбирать отсеки с более высокими температурами по сравнению со II группой. Впервые значения избираемых температур молоди из I группы приблизились к значениям ОИТ уже на 6-е сутки, у II группы ‒ только на 8-е. Стабилизация значений избираемых температур, то есть достижение зоны ОИТ произошло в обеих группах фактически одновременно (разница в одни сутки) и их различие составило 0,6 °С (группа I – 24,2 °С и группа II – 23,6 °С).
Как только молодь окуня обеих групп достигла уровня ОИТ, было произведено первое изменение местоположения укрытий в температурном градиенте. Изъятие укрытий из трех ближайших к зоне температурного оптимума отсеков (№ 7, 8 и 9) в лотке с I группой привело к ощутимым изменениям распределения рыб в температурном градиенте (рис. 2). Так, частота посещения рыбами данных отсеков снизилась почти в два раза, при этом в ближайших к ним отсеках с укрытиями рыбы стали наблюдаться гораздо чаще. Спустя некоторое время (трое суток), по мере адаптации рыб к новым условиям распределения укрытий, встречаемость молоди в зоне температурного оптимума повышалась. Так как рыбы стали чаще наблюдаться в 10-м отсеке, то значения среднесуточной избираемой температуры достоверно (p < 0,05) увеличились (25,0 °С). Через четверо суток после этого укрытия убрали еще из двух отсеков (№ 6 и 10), при этом реакция молоди окуня была схожа с продемонстрированной ранее, то есть увеличилась частота встречаемости рыб в отсеках с сохранившимися укрытиями ближайших к зоне ОИТ (рис. 3).
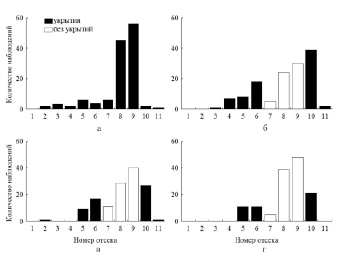
Рис. 2. Распределение молоди I группы в отсеках экспериментальной установки: а – ОИТ (12-е сутки); б – изъятие укрытий из 7, 8 и 9 –го отсеков (13-е сутки); в – 14-е сутки; г – 15-е сутки
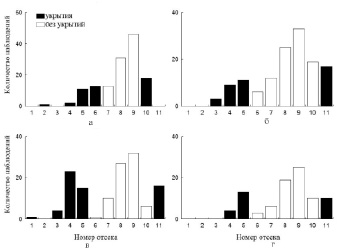
Рис. 3. Распределение молоди I группы в отсеках экспериментальной установки: а – 16-е сутки; б – изъятие укрытий из 6, 7, 8 и 9-го отсеков (17-е сутки); в – 18-е сутки; г – 20-е сутки
В лотке с особями II группы добавление укрытий в отсеки, равноудаленные от температурного оптимума (№ 6 и 10), также внесло определенные коррективы в поведение рыб (рис. 4). Во-первых, рыбы заметнее реагировали на вмешательство в экспериментальную среду. Большая часть из них спустилась в самые «холодные» отсеки и находилась там длительное время. Вследствие этого значения среднесуточной избираемой температуры достоверно (p < 0,05) снизились (21,5 °С). Во-вторых, частота встречаемости молоди в отсеках с добавленными укрытиями (№ 6 и 10) заметно выросла. Дальнейшее перемещение укрытий в 5 и 11 отсеки вызвало схожий эффект (рис. 5).
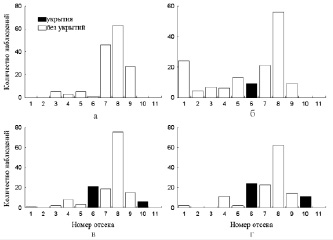
Рис. 4. Распределение молоди II группы в отсеках экспериментальной установки: а – ОИТ (12-е сутки); б – добавлении укрытий в 6-й и 10-й отсеки (13-е сутки); в – 14-е сутки; г – 15-е сутки
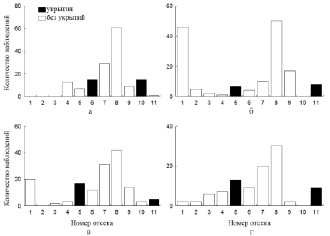
Рис. 5. Распределение молоди II группы в отсеках экспериментальной установки: а – 16-е сутки.; б – перемещение укрытий в 5-ый и 11-ый отсеки (17-е сутки); в – 18-е сутки; г – 20-е сутки
Полученные данные свидетельствуют о том, что молодь окуня в присутствии укрытий способна быстрее осваивать термоградиентную экспериментальную среду. Переход рыб I группы в температуры близкие к оптимальным, происходил заметно быстрее. В среде с малым количеством укрытий у молоди наблюдалась более выраженная стрессорная реакция на вмешательство в экспериментальное пространство (перестановка укрытий). При этом наблюдалось резкое снижение значений избираемых температур в течение суток с момента воздействия. В то время как у молоди I группы при изъятии укрытий из отсеков с температурами, близкими к оптимуму, такой реакции не наблюдалось, и значения избираемых температур не претерпевали сильных изменений. Следовательно, наличие в среде множества укрытий повышает устойчивость молоди к стрессорным воздействиям, позволяя лучше использовать температурный фактор. Следует также отметить, что у рыб обеих групп наблюдался процесс адаптации к смене условий экспериментальной среды. Распределение рыб, претерпевающее изменения после вмешательства (перераспределения укрытий), имело тенденции возвращаться к прежнему. Другими словами, спустя некоторое время частота встречаемости молоди в отсеках со значениями температур, близких к установленному ранее оптимуму, снова повышалась. В то же время ближайшие к зоне ОИТ отсеки с укрытиями сохраняли высокую посещаемость рыбами обеих групп. Как правило, там постоянно находилось от одной до нескольких особей.
К сожалению, литературные данные, рассматривающие влияние укрытий на терморегуляционное поведение рыб, фактически отсутствуют. В единственной найденной нами работе было показано, что малоротый окунь Micropterus dolomieu при наличии в экспериментальной среде укрытий проводил около них большую часть времени, даже если они были расположены вне зоны температурного оптимума (Bevelhimer, 1996). К сожалению, продолжительность нахождения укрытий в той или иной зоне температурного градиента составляла одни сутки, что не позволило отследить дальнейшие тенденции в поведении рыб.
В естественной среде молодь рыб часто использует в качестве укрытий заросли макрофитов, расположенных на прогреваемых участках литорали. При этом молодь окуня способна хорошо использовать пищевые ресурсы, ассоциированные с зарослями растений (крупные виды беспозвоночных) (Persson, 1993). Таким образом, для молоди окуня на заросших прибрежных биотопах создаются хорошие условия для ускоренного роста вследствие оптимизации пищевого и терморегуляционного поведения. В то же время заросли макрофитов предоставляют множество убежищ для защиты молоди рыб от хищников. Однако, как было показано ранее, молодь окуня тяготеет к зарослям растительности как в присутствии, так и отсутствии хищников (Christensen and Persson, 1993).
Заключение
Резюмируя полученные результаты, можно сказать, что при взаимодействии таких факторов внешней среды, как температура и физическая структура местообитаний, первая будет оказывать доминирующее воздействие на встречаемость молоди окуня на тех или иных участках водоемов. В то же время, в присутствии укрытий создаются более благоприятные условия для реализации температурных предпочтений рыб, так как снижаются стрессорные воздействия, вызванные другими факторами, например, возможным присутствием хищников. При этом на заросших растительностью мелководьях, предоставляющих массу укрытий, высокую концентрацию кормовых объектов и повышенные значения температуры воды, создаются прекрасные условия для нагула молоди окуня.
Работа выполнена при финансовой поддержке гранта РФФИ 12-04-31285 мол_а.
Рецензенты:
Герасимов Ю.В., д.б.н., зав. лабораторией экологии рыб, ФГБУН «Институт биологии внутренних вод им. И. Д. Папанина Российской академии наук», пос. Борок;
Терещенко В.Г., д.б.н., с.н.с., гл.н.с. лаборатории эволюционной экологии, ФГБУН «Институт биологии внутренних вод им. И.Д. Папанина Российской академии наук», пос. Борок.
Работа поступила в редакцию 29.11.2013.
Библиографическая ссылка
Смирнов А.К., Смирнова Е.С. ЗАВИСИМОСТЬ ТЕМПЕРАТУРЫ, ПРЕДПОЧИТАЕМОЙ МОЛОДЬЮ ОКУНЯ (Perca fluviatilis L.), ОТ НАЛИЧИЯ УКРЫТИЙ В ЭКСПЕРИМЕНТАЛЬНОЙ СРЕДЕ // Фундаментальные исследования. 2013. № 10-13. С. 2882-2886;URL: https://fundamental-research.ru/ru/article/view?id=32928 (дата обращения: 25.07.2026).