



Злаки (Poaceae) благодаря высокому уровню адаптивности составляют основу большинства лугов всех ботанико-географических зон [8].
На современном этапе особенности отрастания злаков исследованы недостаточно. Изучены связи между биологией отдельных видов и их отрастанием [6; 9], сроками и высотой дефолиации и отрастанием [6], уровнем запасных веществ и отрастанием [5; 6] и т.д. Большой вклад в изучение этой проблемы внесли исследования, проведенные под руководством И.Г. Серебрякова, в которых проанализировано возобновление ряда фестукоидов в зависимости от их побегообразования и ритма развития в различных зонах страны, а также установлена связь между высотой среза, сроками отчуждения и внесением удобрений, с одной стороны, формированием отавы с другой [3]. В ряде отечественных и зарубежных исследований [2; 3; 10] проанализированы общие вопросы отрастания растений. По характеру формирования отавы Т.А. Работнов [4] выделил четыре типа отрастания травянистых растений:
- продолжение роста укороченных побегов;
- продолжение роста удлиненных побегов, если точка роста оказалась выше уровня отчуждения;
- продолжение роста удлиненных побегов в результате образования одного или нескольких побегов из почек в пазухах листьев «пенька», оставшегося после срезания;
- образование новых побегов из почек, расположенных у основания надземных побегов и на подземных органах.
Особенности отрастания бореальных злаков изучены еще также недостаточно. В связи с этим особый интерес представляет исследование отрастания этой группы злаков, характеризующейся многообразием биоморф и составляющей во многих районах основную хозяйственно-ботаническую группу в урожае пастбищ [8].
Продуктивность злаков изучена хорошо, что связано с необходимостью решать практические вопросы сельского хозяйства. Однако мало данных, характеризующих продуктивность и структуру отдельных видов в динамике их развития и по сезонам года [2]. Недостаточно освещены в литературе вопросы побегообразования злаков, их возобновления, отрастания, ритма развития и формирования урожая и некоторые другие, имеющие определенное значение при организации научно обоснованного пастбищного хозяйства. Трудно назвать работы, в которых бы излагались вопросы биоморфологии бореальных злаков во взаимосвязи с их продуктивностью, сезонностью развития и т.д. Между тем подобные исследования имеют большое значение для понимания общих закономерностей развития фестукоидов, их специфичности в сравнении с хорошо изученными паникоидами, а также для решения практических задач по созданию высокопродуктивных долголетних пастбищ и некоторых других вопросов сельского хозяйства этой зоны. Несмотря на большую практическую значимость отрастания бореальных многолетних кормовых злаков, эти вопросы также не нашли в литературе должного отражения [4; 7; 8; 10].
Целью нашей работы является изучение отрастания бореальных многолетних кормовых злаков – отрастание различных побегов и группировка почек.
В настоящей работе излагаются результаты наших исследований отрастания бореальных злаков. Для эксперимента использовались важнейшие в хозяйственном отношении кормовые злаки. В своих исследованиях мы стремились сосредоточить внимание на основных вопросах, часто опуская, на наш взгляд, второстепенные или уже в какой-то степени разработанные и освещенные в работах других авторов. Отрастание злаков изучали на территории Республики Мордовия в 2010–12 гг. Особенности ухода на опытных участках определялись задачами исследований. При изучении отавности отдельных видов их травостой срезали в оптимальные периоды развития с последующим проведением наблюдений за формированием урожая. Для каждого вида отмечали специфику отрастания отдельных побегов. Через определенное время после отчуждения выкапывали растения для камеральной обработки, где подсчитывали число появившихся побегов и отмечали почки, из которых они образовались. Расчет новых структур проводили на каждые 100 отросших побегов в 5–6-кратной повторности. По некоторым видам определяли суточный прирост побегов отавы и вели наблюдения за их развитием.
Отчуждение надземной массы злаков производится тогда, когда продуктивность их травостоя достаточно высокая. По времени это совпадает с фазой выхода в трубку или началом колошения. После отчуждения травостоя разные побеги злаков отрастают неодинаково. Первыми отрастают укороченные побеги. Например, у Lolium perenne L. (рис. 1), Dactylis glomerata L. (рис. 2) и т.д. спустя час после среза можно обнаружить удлинение срезанных влагалищ укороченных побегов. Скорость прироста этих побегов у всех видов очень высокая и в сутки может доходить до 7–9 см. Наибольшим приростом выделяются рыхлодерновинные формы Lolium perenne и Festuca pratensis Huds.
Между злаками наблюдаются большие различия по интенсивности отрастания после скашивания надземной массы в Мордовии.
Неодинаково отрастают также сорта отдельных видов (рис. 3, 4). Отрастание укороченных побегов происходит за счет продолжения роста надрезанных листовых пластинок или влагалищ и образования нетронутым апексом новых фитомеров. Из срезанных побегов у быстро вегетирующих злаков обычно формируются удлиненные структуры. Превращение вегетативных побегов в генеративные обусловлено переходом к этим структурам доминирующей роли, что подтверждается их интенсивным кущением сразу после отчуждения.
Апогеотропные удлиненные побеги после срезания начинают отрастать после укороченных. Возобновляются они двумя путями: пробуждением спящих почек зоны кущения и покоящихся почек надземных удлиненных фитомеров, расположенных ниже уровня среза. Отрастание надземными боковыми почками происходит через 2–5, а подземными – через 5–7 дней после среза. Быстрее отрастают апогеотропные побеги безрозеточных злаков. После срезания надземной части злаков скорость прироста побегов, образующихся из разных почек, неодинакова. Например, в осенний период 2010/2011 гг. в Ичалковском районе в Мордовии у Bromopsis inermis (Leyss.) Holub побеги из апикальных почек прирастали в высоту в среднем на 4,1, из боковых надземных − на 2,8, из почек зоны кущения – на 4, а из почек корневищ – на 3,9 см/сутки.
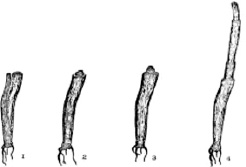
Рис. 1. Отрастание укороченных побегов Lolium perenne: 1 – в момент среза; 2 – через час после среза в послеполуденные часы; 3 – через час после среза в утренние часы; 4 – через 24 часа после среза
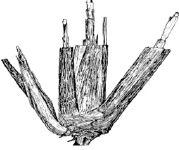
Рис. 2. Отрастание укороченных побегов Dactylis glomerata через 3 часа после среза
Рис. 3. Отрастание укороченных побегов различных сортов Bromopsis inermis после скашивания в Мордовии (2012 г.): –––––– – сорт «Пензенский-1»; – – – – – сорт «Дединовский-3»;  – сорт «Моршанский-312»;
– сорт «Моршанский-312»;  – сорт «Моршанский-760»;
– сорт «Моршанский-760»;  – сорт «Моршанец
– сорт «Моршанец
Рис. 4. Отрастание укороченных побегов различных сортов Phleum pratense L. после скашивания в Мордовии (2012 г.):–––––– – сорт «Вита-1»; – – – – – сорт «Майская-1»
Корневищно-столоновидные побеги после среза растут быстрее, расширяя сферу своего влияния за счет удлинения осевого и образования боковых побегов. Переход их боковых почек в рост почти совпадает по времени с отрастанием укороченных побегов. Корневища реагируют на отчуждение следующим образом: у корневищно-рыхлодерновинных злаков (Bromopsis inermis и др.) верхушечная почка побега принимает апогеотропное направление роста, у корневищных и корневищно-столонообразующих (Elytrigia repens (L.) Nevski, Agrostis stolonifera L.) надземные побеги формируются латеральными и апикальными почками корневищ весеннего происхождения, расположенных ближе к поверхности почвы, на 7–12-й день после среза.
Разнообразие типов побегов в составе особей бореальных злаков обусловило формирование ими большого количества почек, отличающихся рядом особенностей. Образование разнотипных побегов в пределах особи и формирование значительного количества почек возобновления, отличающихся степенью развития, обеспечивает большинству бореальных злаков быстрое отрастание, относительно равномерное накопление урожая в течение вегетации и полицикличность развития. На основе многолетних наблюдений за отрастанием фестукоидов все многообразие почек, участвующих в формировании отавы после отчуждения надземной массы, мы делим на 5 групп, сформированных разными побегами (рис. 5), характеризующимися определенной спецификой развития:
1) апикальные открытые растущие вегетативные почки надземных апо- и диагеотропных побегов (вегетативных укороченных, удлиненных) с невысокой емкостью и расположенных ниже уровня среза. После отчуждения травостоя отрастание идет за счет продолжения удлинения листьев, не закончивших рост к этому времени, а также формирования в конусе нарастания новых листьев. Эта группа включает первые два типа отрастания по Т.А. Работнову [4];
2) боковые закрытые зрелые покоящиеся или растущие почки удлиненных фитомеров надземных диагеотропных побегов (столонов, корневищно-столоновидных), отличающиеся высокой емкостью; почки этой группы быстро переходят в рост и образуют новые структуры;
3) боковые закрытые зрелые или незрелые почки удлиненных фитомеров надземных апогеотропных побегов (генеративных, скрытогенеративных и удлиненных вегетативных), расположенные ниже уровня среза, раскрывающиеся после отчуждения травостоя и характеризующиеся невысокой емкостью. Эта группа включает третий тип отрастания по Т.А. Работнову [4];
4) боковые закрытые зрелые или незрелые почки сближенных фитомеров зоны кущения удлиненных и укороченных побегов, выделяющиеся относительно низкой емкостью;
5) открытые или закрытые почки подземных корневищ (апикальные закрытые зрелые растущие и боковые закрытые зрелые покоящиеся), отличающиеся высокой емкостью, особенно апикальные. Почки четвертой и пятой групп составляют четвертый тип отрастания по Т.А. Работнову [4]. Первые три группы почек расположены в надземной части, две последние – в почве.
Выделенные группы почек отличаются размерами, массой, а также биохимическим составом. Мы определили массу почек у ряда злаков следующим образом: у выкопанных особей каждого вида срезали почки, очищали от чешуй и помещали в каждый пакетик по десять штук в соответствии с группой. По каждой группе брали 10 пакетиков (100 почек). После высушивания определяли массу отдельных почек (табл. 1). В пределах особи почки отдельных видов заметно различаются по своей массе. Наибольшей массой отличаются закрытые растущие почки диагеотропных побегов и апикальных почек корневищ. Заметно выделяются своей массой «луковички» зоны кущения, корневищ и апогеотропных побегов у Calamagrostis epigeios (L.) Roth, в несколько раз превосходя аналогичные почки других видов. Заметно различаются выделенные группы почек и по своей емкости. Этот показатель мы определяли в Мордовии летом и весной (злаки выращены на опытном участке «Республиканского центра дополнительного образования детей» в г. Саранске. Отбирали по 10–12 типичных почек и делали срезы (2–3 почки резали на микротоме, остальные – вручную), которые просматривали под микроскопом и подсчитывали количество листовых образований или их зачатков.
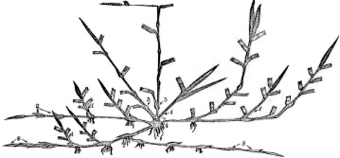
Рис. 5. Схема размещения почек возобновления у бореальных злаков:1–5 – группы почек
Таблица 1
Масса различных групп почек некоторых злаков (Мордовия, 2012)
|
Вид |
Масса почек по группам, мг СВ/почка |
|||||
|
1-я |
2-я |
3-я |
4-я |
5-я |
||
|
апикальные |
латеральные |
|||||
|
Poa pratensis L. s. l. |
0,25 ± 0,07 |
3,41 ± 0,12 |
0,18 ± 0,01 |
2,11 ± 0,14 |
8,12 ± 0,11 |
0,20 ± 0,01 |
|
Bromopsis inermis |
1,52 ± 0,06 |
─ |
11,75 ± 0,53 |
58,40 ± 1,74 |
22,35 ± 0,42 |
21,67 ± 0,70 |
|
Dactylis glomerata |
1,71 ± 0,06 |
─ |
2,16 ± 0,08 |
3,57 ± 0,13 |
13,64 ± 0,18 |
1,94 ± 0,04 |
Следует отметить высокую вариацию емкости в пределах отдельных групп почек (табл. 2). Емкость почек не является стабильной для отдельных видов. По нашим наблюдениям, зрелость почки (переход в растущее состояние) при разных условиях вегетации растений и в зависимости от их возраста различная. Наибольшей емкостью отличаются открытые почки вегетативных апогеотропных побегов короткодневных злаков при их вегетации на длинном дне. Например, апикальные растущие почки Sorghum sudanense в среднем имели свыше 10 зачатков. Большой емкостью отличаются «луковички» (до 15 зачатков), формирующиеся на столонах корневищно-столонообразующих и в зоне кущения и на удлиненных побегах корневищно-рыхлодерновинных злаков. «Луковички» выделяются крупностью и хорошо защищены от пересыхания большим количеством (до 20 и больше) плотных чешуй. Следует указать на различия емкости почек в летний и осенний периоды: осенью, как правило, емкость соответствующих почек на 2–3 метамера меньше, чем летом, что связано, очевидно, с замедлением митоза и общим изменением энергетико-вещественного обмена в растении и переходом почек в состояние покоя. Результаты наших исследований емкости почек бореальных злаков значительно пополняют информацию о злаках. В частности, на незначительное количество примордиев на конусах нарастания маисовых и сорговых указывает Sharman [11] и лесных столонообразующих злаков – Т.И. Серебрякова [8]. Некоторые различия между почками выделенных групп наблюдаются также и в биохимическом составе, в частности, в структуре белков. Активно вегетирующие апикальные почки надземных и поземных побегов отличаются высоким содержанием легкорастворимых белков (до 4 мг/г на СВ и больше), тогда как боковые спящие выделяются низким содержанием этой фракции белков, особенно альбуминов.
Таблица 2
Емкость почек некоторых злаков (Мордовия, 2010–11)
|
Вид |
Тип побега |
Группа почек |
Емкость почек |
Тип почки |
|
|
летом |
осенью |
||||
|
Bromopsis inermis |
Апогеотропный |
1 |
3,3 ± 0,50 |
2,3 ± 0,07 |
ор |
|
Диагеотропный |
2 |
8,5 ± 0,50 |
6,0 ± 0,38 |
зп |
|
|
Апогеотропный |
3 |
2,3 ± 0,07 |
2,6 ± 0,28 |
зп |
|
|
4 |
10,5 ± 0,07 |
7,6 ± 0,28 |
зп |
||
|
Корневище, апекс |
5 |
10,4 ± 0,07 |
5,5 ± 0,75 |
зр |
|
|
Боковые |
5 |
7,9 ± 0,32 |
5,6 ± 0,27 |
зп |
|
|
Dactylis glomerata |
Апогеотропный |
1 |
6,3 ± 0,20 |
2,3 ± 0,17 |
ор |
|
2 |
─ |
─ |
─ |
||
|
3 |
4,5 ± 0,34 |
2,3 ± 0,17 |
зп |
||
|
4 |
5,6 ± 0,34 |
3,6 ± 0,33 |
зп |
||
|
5 |
─ |
─ |
─ |
||
|
Lolium multiflorum |
Апогеотропный |
1 |
4,4 ± 0,53 |
3,5 ± 0,28 |
ор |
|
2 |
─ |
─ |
─ |
||
|
3 |
2,2 ± 0,18 |
2,3 ± 0,19 |
зп |
||
|
4 |
5,6 ± 0,28 |
2,8 ± 0,17 |
зп |
||
|
Calamagrostis epigeios |
Апогеотропный |
1 |
6,3 ± 0,27 |
5,0 ± 0,47 |
ор |
|
2 |
─ |
─ |
─ |
||
|
3 |
5,6 ± 0,33 |
4,8 ± 0,21 |
зр |
||
|
«Луковичка» |
4 |
12,4 ± 0,30 |
11,0 ± 0,46 |
зп |
|
|
Корневище |
5 |
9,7 ± 0,33 |
9,6 ± 0,55 |
зр |
|
|
Poa pratensis |
Апогеотропный |
1 |
4,8 ± 0,33 |
3,7 ± 0,30 |
ор |
|
2 |
─ |
─ |
─ |
||
|
3 |
4,4 ± 0,29 |
4,3 ± 0,19 |
зр |
||
|
4 |
5,3 ± 0,19 |
4,8 ± 0,40 |
зп |
||
|
Корневище, апекс |
5 |
9,6 ± 0,29 |
7,6 ± 0,29 |
зр |
|
|
Боковые |
5 |
8,3 ± 0,14 |
5,8 ± 0,34 |
зп |
|
|
Sorghum sudanense |
Апогеотропный |
1 |
11,4 ± 0,22 |
5,3 ± 0,25 |
ор |
Примечание. ор − открытая растущая; зр − закрытая растущая; зп – закрытая покоящаяся.
Проведенный анализ ряда показателей различных почек (биометрический и биохимический) свидетельствует о значительных различиях в их характеристике. Отмеченные расхождения между почками коррелируют с их потенциальными возможностями в формировании побеговых структур. Это подтверждается также данными наших исследований за отрастанием костреца безостого (Bromopsis inermis) сорта «Пензенский-1» в весенний и летний периоды года в Мордовии. Растения срезали на высоте 10 см, спустя 30 дней брали произвольно по 25 побегов, образованных разными почками, и производили их анализ (табл. 3).
Образованные разными группами почек побеги заметно различаются по характеру роста (прирост, масса и т.д.). Мощным накоплением урожая в летний период выделялись побеги подземного происхождения, а весной – побеги, образованные апикальными почками, а также почками корневищ и зоны кущения. Отмеченные особенности в формировании отавы при участии различных групп почек имеют большое практическое значение, указывая на необходимость тщательного подхода к вопросу об уровне дефолиации пастбищ.
Несмотря на большие различия почек отдельных групп по ряду показателей (масса, емкость, биохимический состав и т.д.), следует отметить известную условность такого деления, что связано с широким варьированием биометрических характеристик почек и их различной степенью развития в пределах отдельных групп.
Таблица 3
Характеристика побегов Bromopsis inermis, образованных почками разных групп
|
Побеги, образованные почками |
Характеристика побегов |
||||
|
высота, см |
диаметр, см |
масса, г/СВ |
суточный прирост, см |
тип побега |
|
|
Лето 2011 г. |
|||||
|
Боковыми надземными |
134 ± 5,3 |
0,9 |
11,2 ± 0,24 |
4,4 ± 0,04 |
уд. |
|
Зоны кущения |
320 ± 4,3 |
1,4 |
29,7 ± 0,35 |
10,7 ± 0,21 |
уд. |
|
Корневищ |
320 ± 2,8 |
2,0 |
32,5 ± 0,34 |
10,7 ± 0,14 |
уд. |
|
Весна 2011–12 гг. |
|||||
|
Апикальными |
123 ± 2,5 |
1,0 |
11,4 ± 0,30 |
4,0 ± 0,08 |
уд. |
|
Боковыми надземными |
84 ± 1,3 |
0,5 |
1,3 ± 0,05 |
2,8 ± 0,04 |
уд. |
|
Зоны кущения |
120 ± 3,1 |
0,8 |
4,2 ± 0,08 |
4,0 ± 0,06 |
ук. |
|
Корневищ |
117 ± 1,8 |
0,9 |
4,3 ± 0,06 |
3,9 ± 0,06 |
ук. |
Примечание: уд. – удлиненные; ук. – укороченные.
В подтверждение этого приведем некоторые данные по характеристике боковых почек различных частей удлиненных вегетативных побегов (среднее из 25 замеров) Phragmites australis (Cav.) Trin. ex Steud, который образует заросли на сырых местах и по берегам водоемов в Мордовии (табл. 4).
Таблица 4
Характеристика почек удлиненного вегетативного побегаPhragmites australis (Саранск, 2012)
|
Номер почек, снизу вверх |
Характеристика почек |
Примечание |
|||
|
длина, мм |
диаметр, мм |
масса, мг /СВ |
форма |
||
|
1 |
11 |
7 |
0,27 ± 0,005 |
Округлая |
Первые 5 метамеров составляют зону кущения, 3–5-я почки тронулись в рост |
|
2 |
13 |
7 |
0,36 ± 0,008 |
-«- |
|
|
3 |
19 |
11 |
0,40 ± 0,008 |
-«- |
|
|
4 |
14 |
12 |
0,46 ± 0,006 |
-«- |
|
|
5 |
14 |
11 |
0,16 ± 0,004 |
-«- |
|
|
6 |
6 |
6 |
0,09 ± 0,004 |
-«- |
Почки 6–10-го фитомеров находятся в состоянии покоя (зона покоящихся надземных боковых почек) |
|
7 |
7 |
7 |
0,05 ± 0,002 |
-«- |
|
|
8 |
4 |
4 |
0,09 ± 0,003 |
Округло-плоская |
|
|
9 |
10 |
4 |
0,05 ± 0,003 |
-«- |
|
|
10 |
7 |
3 |
0,02 ± 0,001 |
-«- |
|
|
11 |
8 |
2 |
0,01 ± 0,001 |
-«- |
Почки 11–22-го метамеров находятся в фазе формирования (зона растущих боковых надземных почек) |
|
12 |
6 |
2 |
0,01 ± 0,001 |
-«- |
|
|
13 |
6 |
2 |
0,01 ± 0,001 |
-«- |
|
|
14 |
6 |
2 |
0,01 ± 0,001 |
-«- |
|
|
15 |
5 |
1,5 |
0,05 ± 0,001 |
-«- |
|
|
16 |
5 |
1 |
0,009 ± 0,002 |
-«- |
|
|
17 |
5 |
1 |
0,002 ± 0,0001 |
-«- |
|
|
18 |
4 |
1 |
0,002 ± 0,0001 |
-«- |
|
|
19 |
4 |
1 |
0,002 ± 0,0004 |
-«- |
|
|
20 |
3 |
1 |
0,002 ± 0,0003 |
-«- |
|
|
21 |
3 |
1 |
0,002 ± 0,0001 |
-«- |
|
|
22 |
2 |
1 |
0,002 ± 0,0001 |
-«- |
|
Параметры почек разных метамеров в подземной и надземных частях заметно варьируются, что свидетельствует о разной степени их развития. Наиболее крупными являются почки средних метамеров, которые раньше других переходят в рост.
Следует отметить определенную закономерность в степени развития почек в зоне кущения и в надземной части апогеотропного побега: крайние верхние и нижние почки (самые ранние и поздние) отстают в росте от срединных и нередко останавливаются на этом этапе развития. Средние почки зоны кущения и надземной части образуют боковые структуры в период жизнедеятельности материнского побега. Нижние почки удлиненных фитомеров и некоторые почки зоны кущения остаются в покоящемся состоянии и раскрываются после отчуждения надземной части, причем первыми в рост трогаются почки надземных фитомеров, а затем зоны кущения.
Существует определенная связь между формированием кольца придаточных корней и раскрытием почки соответствующего фитомера. Например, в зоне кущения у Bromopsis inermis почки раскрываются быстрее в том случае, если кольцо придаточных корней вокруг их узла отсутствует. Побеги, у которых все подземные узлы образовали придаточные корни, после отчуждения надземной части отрастают, как правило, не почками зоны кущения, а почками корневищ.
Таким образом, проведенный анализ отрастания злаков показывает, что формирование отавы после отчуждения травостоя обусловлено их биологическими особенностями и прежде всего структурой формируемых особей, а также размещением почек возобновления и уровнем развития последних.
Рецензенты:
Любарский Е.Л., д.б.н., профессор кафедры ботаники биолого-почвенного факультета ФГБОУ ВПО «Казанский (Приволжский) федеральный университет, г. Казань;
Силаева Т.Б., д.б.н., профессор кафедры ботаники и физиологии растений ФГБОУ ВПО «Мордовский государственный исследовательский университет им. Н.П. Огарева, г. Саранск.
Работа поступила в редакцию 10.01.2013.
Библиографическая ссылка
Горчакова А.Ю. БОРЕАЛЬНЫЕ ЗЛАКИ – НЕКОТОРЫЕ ОСОБЕННОСТИ ОТРАСТАНИЯ // Фундаментальные исследования. 2013. № 1-3. С. 560-567;URL: https://fundamental-research.ru/ru/article/view?id=30985 (дата обращения: 20.06.2026).