



Прибрежная зона моря является первым уровнем, на который поступает основное количество загрязняющих веществ от источников, расположенных на его берегах. В этом плане, морские водоросли, являясь первичными продуцентами органического вещества, играют решающую роль в функционировании всей экосистемы водоёма и отдельных ее компонентов. Макрофитобентос выполняет роль мощного фильтра различных загрязняющих веществ, поступающих в его пределы, участвует в потреблении, распределении, переработке этих веществ и, по сути, обеспечении питания других структурных уровней шельфовой зоны моря.
Цемесская бухта по масштабам портово-промышленной и транспортной деятельности и, следовательно, по совокупности антропогенных воздействий на морскую среду не имеет себе равных в Азово-Черноморском регионе России. Существенный вклад в загрязнение прибрежной зоны восточного побережья бухты вносят нефтепродукты, попадающие в море в составе сточных вод перевалочной нефтебазы «Шесхарис» (далее по тексту – ПНБ «Шесхарис»). Начиная с 1963 и до 1977 г. сброс частично очищенных нефтесодержащих сточных вод предприятия осуществлялся из системы прудов-отстойников через подводный дюкер. В последующие 35 лет сточные воды отводятся в море в другом районе и только через глубоководный выпуск (удаление от берега 600 м). Расстояние от оголовка нового выпуска сточных вод предприятия до старого выпуска из прудов-отстойников составляет около 1 км.
Макрофиты, являясь прикрепленными организмами моря, представляют главную мишень воздействия нефтепродуктов, они особенно остро реагируют на любые изменения качества водной среды. Изучение видового состава и функционирования прибрежных водорослевых сообществ в условиях нефтяного загрязнения моря тесно связано с познанием процессов самоочищения водной среды и скорости восстановления фитоценозов.
Актуальность этой проблемы, исходя из требований российского законодательства в области охраны окружающей среды, а также с точки зрения решения практических задач гидротехнического и рекреационного строительства, очевидна.
Макрофитобентос Цемесской бухты относительно хорошо изучен. Большая часть работ посвящена изучению водорослей в летний период и в наиболее экологически напряженных районах (порт и прилегающая акватория). На сегодняшний день отсутствуют данные о современном состоянии макрофитобентоса в районе старого выпуска нефтесодержащих сточных вод ПНБ «Шесхарис» из системы прудов-отстойников, а по району нового глубоководного выпуска имеются лишь разрозненные данные разных лет исследований, выполненные в летний период года.
Цель данной работы – исследование эколого-таксономического разнообразия осеннего макрофитобентоса в районах старого и нового выпусков нефтесодержащих сточных вод ПНБ «Шесхарис».
Материал и методы исследований
Объектом исследований являлись водорослевые бентосные сообщества районов старого и нового выпусков нефтесодержащих сточных вод ПНБ «Шесхарис» (рис. 1).
Работы выполнялись на участке шельфа восточного побережья Цемесской бухты от нефтегавани до прудов-отстойников. Отбор качественных проб осуществлялся в конце сентября – начале октября 2013 г. по общепринятой гидроботанической методике (Калугина, 1969) с помощью водолаза с берега (глубины до 3 м) и малых плавсредств (глубины до 10 м). Всего было отобрано и обработано 32 качественные пробы водорослей. Видовой состав макрофитов определяли по «Определителю зеленых, бурых и красных водорослей южных морей СССР» [1] с учетом современных номенклатурных изменений [2–4]. Водоросли определены до вида. Для сравнительной оценки видовой структуры макрофитобентоса в районах наблюдений применены коэффициенты встречаемости (R), флористического сходства (КJ), флористический (Рфлор), предложенный Д.Т. Ченеем [7–9].
Результаты исследований и их обсуждение
На участке исследований прибрежная зона моря характеризуется выраженным микрорельефом дна с резко нарастающими глубинами. От уреза воды до глубины 5–7 м дно слагается галечниково-валунным материалом, пересечено скальными грядами, в складках – накопления мелкого гравийного материала с песком и признаками заиления. Глубже микрорельеф сглаживается, уступая место алеврито-пелитовым илам. На галечниках до глубины 0,3–0,5 м макроводоросли не обнаружены, что связано с высокой прибойностью и характером сложения донных грунтов.

Рис. 1. Карта-схема расположения районов исследований макрофитобентоса в Цемесской бухте
Проведенные исследования в двух районах (нового и старого выпусков) позволили обнаружить 57 видов макрофитов, относящихся к 33 родам, 21 семейству, 17 порядкам отделов Chlorophyta, Phaeophytа и Rhodophyta (таблица). По разнообразию таксономического состава водорослей доминируют виды отдела Rhodophyta, составляя более 57 % общего числа видов и родов, 48 % семейств, 41 % порядков. Наиболее многочисленный в видовом отношении порядок Ceramiales (3 семейства и 9 родов) включает 35,1 % общего числа видов макрофитобентоса. Основу видового разнообразия составляют семейства Ceramiaceae и Rhodomelaceae, на долю которых в сумме приходится 57,6 % общего числа видов данного отдела водорослей.
Видовой состав макрофитобентоса в районах выпусков нефтесодержащих сточных вод ПНБ «Шесхарис» (28.09. – 04.10.2013 г.)
|
№ п/п |
Отдел, вид |
Экологические показатели* |
Район |
|
|
Новый выпуск |
Старый выпуск |
|||
|
1 |
2 |
3 |
4 |
5 |
|
Phaeophyta |
||||
|
1. |
Myrionema balticum (Reinke) Foslie |
М |
– |
+ |
|
2. |
Ectocarpus confervoides (Roth) Le Jolis |
М |
– |
+ |
|
3. |
Entonema oligosporum (Strömf.) |
М |
+ |
+ |
|
4. |
Nereia filiformis (J.Ag.) Zanard. |
О |
– |
+ |
|
5. |
Sphacelaria cirrosa (Roth) Ag. |
М |
+ |
+ |
|
6. |
Cladostephus verticillatus (Lightf.) |
О |
+ |
+ |
|
7. |
Cystoseira barbata (Good. et Wood.) Ag. |
М |
+ |
+ |
|
8. |
C. crinita Bory |
О |
+ |
+ |
|
9. |
Padina pavonia (L.) Gaill. |
О |
+ |
– |
|
Chlorophyta |
||||
|
1. |
Acrosiphonia centralis (Lyngb.) Kjellm. |
О |
– |
+ |
|
2. |
Enteromorpha prolifera (O. Müll.) J. Ag. |
М |
+ |
+ |
|
3. |
E. linza (L.) J. Ag. |
М |
+ |
– |
|
4. |
E. intestinalis (L.) Link. |
П |
+ |
+ |
|
5. |
E. flexuosa (Wulf.) J. Ag. |
М |
+ |
– |
|
6. |
Chaetomorpha aёrea (Dillw.) Kütz. |
М |
+ |
+ |
|
7. |
Cladophora echinus (Bias.) Kütz. |
О |
– |
+ |
|
8. |
C. albida (Huds.) Kütz. |
П |
+ |
+ |
|
9. |
C. laetevirens (Dillw.) Kütz. |
П |
+ |
+ |
|
10. |
C. vadorum (Aresch.) Kütz. |
М |
+ |
+ |
|
11. |
C. liniformis Kütz. |
П |
+ |
– |
|
12. |
C. sericea (Huds.) Kütz. |
М |
– |
+ |
|
13. |
C. dalmatica Kütz. |
О |
– |
+ |
|
14. |
Cladophoropsis membranacea (Ag.) Börg. |
П |
+ |
+ |
|
15. |
Codium vermilara (Olivi) Delle Chiaje |
О |
+ |
+ |
|
Rhodophyta |
||||
|
1. |
Asterocytis ramosa (Thw.) Gobi |
П |
– |
+ |
|
2. |
Goniotrichum elegans (Chauv.) Zanard. |
М |
– |
+ |
|
3. |
Erythrotrichia carnea (Dillw.) J.Ag. |
П |
– |
+ |
|
4. |
Kylinia secundata (Lyngb.) Papenf. |
М |
+ |
+ |
|
5. |
K. virgatula (Harv.) Papenf. |
М |
+ |
– |
|
6. |
Gelidium crinale (Turn.) Lamour. |
О |
+ |
+ |
|
7. |
G. latifolium (Grev.) Born. et Thur. |
М |
+ |
+ |
|
8. |
Melobesia farinosa Lamour. |
? |
+ |
+ |
|
9. |
Corallina mediterranea Aresch. |
О |
+ |
+ |
|
10. |
C. granifera Ell. et Soland. |
О |
– |
+ |
|
11. |
Jania rubens (L.) Lamour. |
О |
+ |
– |
|
12. |
Gracilaria verrucosa (Huds.) Papenf. |
О |
– |
+ |
|
1 |
2 |
3 |
4 |
5 |
|
13. |
Phyllophora nervosa (DC.) Grev. |
О |
+ |
+ |
|
14. |
Antithamnion plumula (Ell.) Thur. |
П |
– |
+ |
|
15. |
Ceramium ciliatum (Ell.) Ducl. |
О |
+ |
+ |
|
16. |
C. rubrum (Huds.) Ag. |
П |
+ |
+ |
|
17. |
C. elegans Ducl. |
П |
+ |
+ |
|
18. |
C. tenuissimum (Lyngb.) J.Ag. |
М |
– |
+ |
|
19. |
C. diaphanum (Lightf.) Roth. |
П |
– |
+ |
|
20. |
Callithamnion corymbosum (J. E. Smith) Lyngb. |
М |
+ |
– |
|
21. |
Spermothamnion strictum (Ag.) Adriss. |
М |
+ |
+ |
|
22. |
Apoglossum ruscifolium (Turn.) J. Ag. |
М |
+ |
+ |
|
23. |
Polysiphonia denudata (Dillw.) Kütz. |
О |
+ |
+ |
|
24. |
P. subulifera (Ag.) Harv. |
М |
+ |
+ |
|
25. |
P. breviarticulata (Ag.) Zanard. |
? |
– |
+ |
|
26. |
P. opaca (Ag.) Zanard. |
М |
+ |
+ |
|
27. |
P. pulvinata Kütz. |
О |
– |
+ |
|
28. |
Lophosiphonia reptabunda (Suhr) Kylin |
М |
– |
+ |
|
29. |
Chondria tenuissima (Good. et Wood) Аg. |
О |
+ |
+ |
|
30. |
Ch. dasyphylla (Wood.) Ag. |
О |
– |
+ |
|
31. |
Laurencia paniculata J. Ag. |
О |
– |
+ |
|
32. |
L. obtusa (Huds.) Lamour. |
М |
+ |
+ |
|
33. |
L. hybrida (DC.) Lenorm. |
О |
– |
+ |
|
Всего |
36 |
50 |
||
Примечание. * – О – олигосапробы, М – мезосапробы, П – полисапробы; ? – не известно.
Количество видов Chlorophyta в 1,7 раза превышает Phaeophytа. Среди Chlorophyta по таксономическому составу доминирует порядок Cladophorales (60 % общего числа видов), у Phaeophytа распределение видов по отдельным надвидовым таксонам примерно одинаковое.
Большинство семейств (57,1 %) и родов (69,7 %) относятся к монотипическим. Высоким видовым разнообразием характеризуются Сladophoraceae, Rhodomelaceae и Ceramiaceae, на их долю приходится до 49 % общего числа зарегистрированных видов водорослей. Наибольшую видовую насыщенность имеет род Cladophora (7 видов), в составе родов Polysiphonia и Ceramium – по 5 видов. Среднее число видов в роде составляет 2,6.
Отличительной особенностью структуры фитоценозов исследованных районов является высокое видовое разнообразие (87,7 общего числа видов) сообществ у старого выпуска. Ведущее положение в структуре имеют Phaeophytа и Rhodophyta (составляют в сумме более 66 % видов). При этом в таксономической структуре Phaeophytа число родов и порядков не зависит от принадлежности сообщества к тому или иному исследованному району. Из Chlorophyta высоким видовым разнообразием отличается Сladophoraceae (66,7 % от общего числа видов отдела).
В районе нового выпуска зарегистрировано на 28 % видов меньше, чем у старого выпуска. Макроводоросли, принадлежащие к порядку Dictyotales, отмечены только в районе нового выпуска. Коэффициент флористического сходства состава выделенных таксонов альгофлор варьирует в интервале 32,7–35,7. По количеству видов наибольшее отличие обнаружено у Rhodophyta. Особенностью родовой структуры альгофлоры района нового выпуска является более низкая видовая насыщенность Polysiphonia, Ceramium и Cladophora и более высокая – Enteromorpha. Сопоставление изменчивости таксономической структуры отделов в каждом из характеризуемых районов показало, что наибольший предел вариаций числа видов прослеживается у порядков Rhodophyta и Chlorophyta, у семейств – Rhodomelaceae, Ceramiaceae и Сladophoraceae.
Малое количество бурых видов водорослей в исследованных районах (15,8 % общего числа видов), являющихся индикаторами чистых вод, определило высокое значение флористического коэффициента Чени (Рфлор = 5,3), что характерно для районов со средней степенью загрязнения морской воды (пределы Рфлор от 3 до 6). Распределение видов по группам сапробности показало превалирование олигосапробов в районе старого выпуска (38 % общего числа видов). Это позволяет сделать вывод, что основным фактором, влияющим на флористический состав макроводорослей в районе нового выпуска, является хроническое нефтяное загрязнение.
В целом макрофитобентос исследованных районов характеризуется выраженной индивидуальностью флористического состава (рис. 2). На это указывает и тот факт, что виды 4 порядков (Chordariales, Sporochnales, Goniotrichales, Bangeales) отмечены только у старого выпуска.
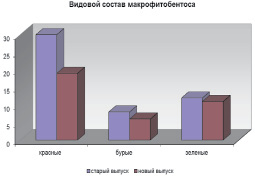
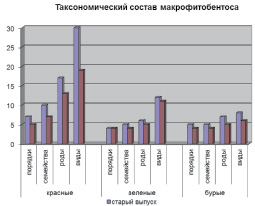
Рис. 2. Флористическая структура макрофитобентоса в районах старого и нового выпусков нефтесодержащих сточных вод ПНБ «Шесхарис»
Анализ распределения видов макроводорослей по величине коэффициента встречаемости показал, что в составе альгофлоры районов наибольшее количество видов (56,1 % общего числа видов) имеют низкий показатель (R = 1 – < 20 %) и относятся к I классу постоянства [5]. Доля видов, отнесенных к наивысшему V классу (R = 81–100 %), составляет всего 15,8 % общего числа видов.
Заключение
Район произрастания макроводорослей, а вернее уровень нефтяного загрязнения вод, определенным образом влияет на основные характеристики его водорослевых сообществ.
Альгофлора исследованных районов старого и нового выпусков нефтесодержащих сточных вод ПНБ «Шесхарис» насчитывает 57 видов макрофитов, относящихся к 33 родам, 21 семейству, 17 порядкам отделов Chlorophyta, Phaeophytа и Rhodophyta. По разнообразию таксономического и видового составов водорослей доминирует Rhodophyta, в его составе – порядок Ceramiales.
Таксономическая и видовая структура фитоценозов изученных районов имеет выраженные качественные различия. В составе макрофитобентоса районов отмечено около половины одинаковых видов. Сходство альгофлор подчеркивается доминированием количественных показателей отделов в последовательности Rhodophyta → Chlorophyta → Phaeophytа. Совпадение видовых таксонов увеличивается в ряду Phaeophytа → Chlorophyta → Rhodophyta.
В районе нового выпуска нефтесодержащих сточных вод таксономическая структура макрофитобентоса упрощенная, что обусловлено хроническим нефтяным загрязнением водной среды. У старого выпуска фитобентос находится в стадии естественных сукцессионных процессов, направленных на восстановление видового разнообразия и структуры сообществ.
Рецензенты:
Туркин В.А., д.т.н., профессор, начальник кафедры «Техносферная безопасность на транспорте», ФГБОУ ВПО «ГМУ им. адм. Ф.Ф. Ушакова», г. Новороссийск;
Страхова Н.А., д.т.н., профессор кафедры «Техносферная безопасность на транспорте», ФГБОУ ВПО «ГМУ им. адм. Ф.Ф. Ушакова», г. Новороссийск.
Библиографическая ссылка
Березенко Н.С. ЭКОЛОГО-ТАКСОНОМИЧЕСКИЙ СОСТАВ МАКРОФИТОБЕНТОСА РАЙОНА ВЫПУСКА НЕФТЕСОДЕРЖАЩИХ СТОЧНЫХ ВОД ПНБ «ШЕСХАРИС» (ЦЕМЕССКАЯ БУХТА, ЧЕРНОЕ МОРЕ) // Фундаментальные исследования. 2015. № 6-2. С. 219-224;URL: https://fundamental-research.ru/ru/article/view?id=38544 (дата обращения: 07.07.2026).