



Матка как гетерогенный, полифункциональный орган имеет многоуровневую систему регуляции, в которой важное значение имеют биогенные амины - катехоламины, серотонин и гистамин [1, 11]. В зависимости от рабочей ситуации биоамины могут выполнять функцию посредников между гормонами и рецепторно-циклазными системами, оказывать адаптирующее и регулирующее действие на клетки, ткани и органы [1, 3-5]. В настоящее время мало изучены экстраорганные элементы биоаминового обеспечения матки – брыжейка, перитонеальная жидкость, хотя именно они играют заметную роль в развитии патологии женской репродуктивной системы [2].
Целью нашей работы явилось обобщение данных литературы и результатов собственных исследований, посвященных участию внеорганных структур (перитонеальная жидкость, кровь, брыжейка матки) в системе регуляции биоаминового обеспечения матки в процессе полового цикла.
В результате проведенных нами микроспектрофлуориметрических исследований перитонеальной жидкости крыс серотонин, катехоламины и гистамин дифференцированы в тучных клетках, макрофагах и жидкостной фазе [7-9]. Определено содержание моноаминов в указанных структурах в различные фазы полового цикла.
Результаты наших исследований подтверждают данные о наличии гистамина и серотонина в тучных клетках [12, 13] и макрофагах [12, 17] перитонеальной жидкости крыс. Часть синтезируемых тучными клетками биоаминов [15], по всей видимости, регулярно выделяется в жидкостную фазу перитонеальной жидкости, что косвенно подтверждает обнаружение нами в ней достоверно значимых концентраций гистамина, серотонина и катехоламинов. Из жидкости моноамины могут захватываться макрофагами.
В последнее время установлена способность перитонеальных макрофагов продуцировать гистамин и серотонин [14, 16] и регулировать активность тучных клеток [18]. Нам не удалось найти литературных данных о способности перитонеальных макрофагов самостоятельно секретировать катехоламины. Согласно предложенной нами рабочей гипотезе о функциональной кооперации элементов внутриорганного комплекса биоаминового обеспечения макрофаги осуществляют секрецию (гистамина и серотонина), поглощение, накопление и, возможно, инактивацию излишков моноаминов, которые в силу тех или иных обстоятельств не утилизировались рабочими структурами [6]. Вероятно, в ряде случаев макрофаги выполняют функцию переносчиков биоаминов между различными участками органа, а в некоторых случаях могут служить дополнительными их источниками.
Основными биоаминпозитивными структурами брыжейки матки являются симпатические нервные волокна, тучные клетки, макрофаги [8, 10]. Микроспектральный анализ флуоресценции нервных волокон брыжейки свидетельствует о наличии в них наряду с катехоламинами и серотонина, что подтверждает данные о его медиаторной функции в периферических отделах вегетативной нервной системы [4, 11]. Наибольшее увеличение содержания серотонина и катехоламинов во всех структурных элементах нервных волокон отмечается в стадии раннего и позднего эструса, а снижение – в ранний диэструс.
Основными структурами симпатических нервных волокон, осуществляющими эффекторную реализацию нейромедиаторов, их синтез, захват и депонирование, являются варикозные расширения, а межварикозные участки осуществляют в основном транспортную роль [4, 5]. Проведенный нами сравнительный анализ уровня изменений одноименных биоаминов в различных структурах нервных волокон в составе ПВС демонстрирует более выраженную динамику серотонина и катехоламинов именно в межварикозных участках [10].
Можно предположить, что нервные волокна брыжейки матки осуществляют транспортировку биогенных аминов в матку в зависимости от потребностей органа в каждый конкретный рабочий момент полового цикла.
Синхронно изменениям содержания катехоламинов и серотонина в нервных волокнах изменяется их уровень в тучных клетках [7, 10]. В настоящее время общепризнанно, что тучные клетки являются посредниками между нервной и гуморальной системами организма с одной стороны и «рабочими» клетками органов с другой. Поглощая невостребованные эффекторными клетками биогенные амины, транспортируя и инактивируя их, тканевые базофилы являются регуляторами биоаминового обмена в органе [5].
Минимальная концентрация серотонина и катехоламинов в макрофагах брыжейки отмечается в ранний и поздний эструс, когда наблюдается пик их уровня в нервных волокнах [10]. Большое содержание моноаминов как в макрофагах перитонеальной жидкости, так и брыжейки матки, по всей видимости, отражает количество их свободных форм в межклеточном пространстве, которые могут быть подвергнуты поглощению макрофагами [5].
Статистически достоверно установлено, что в мазке периферической крови наименьшее содержание исследуемых биоаминов наблюдается в метаэструсе, максимальный уровень катехоламинов и гистамина приходится на поздний эструс, серотонина – на ранний диэструс [9].
На протяжении всего полового цикла нами выявлено сохранение высокой степени положительной линейной корреляции между уровнями серотонина и катехоламинов в крови, перитонеальных и мезентериальных тучных клетках и макрофагах, жидкостной фазе перитонеальной жидкости, нервных волокнах брыжейки в точках зондирования. Данный факт свидетельствует о сильном сопряжении количества биоаминов-антагонистов в исследуемых структурах. Дисбаланс концентраций моноаминов может быть одной из причин патологии органов женской репродуктивной системы.
Установлен ряд значимых хроносопряжений в динамике изменений оценочных параметров исследуемых биоаминпозитивных структур в течение полового цикла
По данным рангового корреляционного анализа Спирмена выявлен ряд достоверных сопряжений изменений в течение полового цикла оценочных параметров биоаминпозитивных структур перитонеальной жидкости, периферической крови, матки и ее брыжейки.
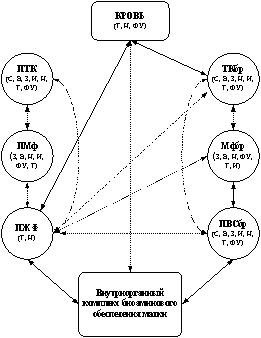
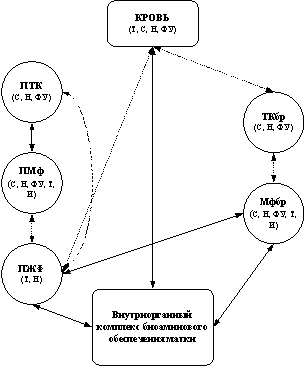
Результаты наших исследований в совокупности с данными литературы позволили выделить своеобразный комплекс экстраорганного биоаминового обеспечения матки, состоящий из нескольких звеньев: 1) перитонеальное, 2) мезентериальное, 3) звено периферической крови. Указанные звенья, различающиеся по генезу и составу, осуществляют синтез, захват, функциональную реализацию, накопление, инактивацию и транспорт к биоаминпозитивным структурам матки серотонина, катехоламинов и гистамина (рис. 1).
 а
а 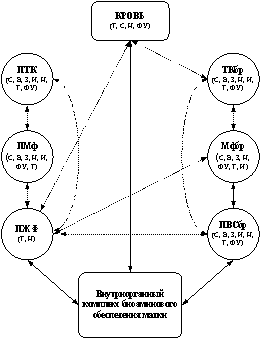 б
б
 в
в
Рис. 1. Схема структурно-функциональной интеграции экстраорганных элементов обеспечения матки катехоламинами (а), серотонином (б) и гистамином (в). Стрелками показаны возможные пути обеспечения моноаминов, пунктирная линия свидетельствует о дискуссионности наличия связи (нулевая гипотеза не отвергается).
ПТК – перитонеальная тучная клетка, ПМф – перитонеальный макрофаг, ПЖФ – жидкостная фаза перитонеальной жидкости, ТКбр – тучная клетка брыжейки матки, Мфбр – макрофаг брыжейки матки, ПВСбр – периваскулярные сплетения брыжейки матки, С – синтез, Н – накопление, Т – транспорт, И – инактивация, ФУ – функциональная утилизация, В – выведение, З – захват
Каждое звено обладает элементами надежности (резервность, избыточность, восполняемость), механизмами получения и обработки информации, каналами реализации полезного действия, а также контроля и коррекции программного результата. Нарушения, возникающие в том или ином звене, могут привести к изменению внутриорганного биоаминового обмена и стать причиной развития патологии матки. Ведущим фактором в этом процессе может быть не столько изменение абсолютных значений уровня моноаминов в микроокружении эффекторных клеток, сколько нарушение их количественного соотношения.
Таким образом, содержание катехоламинов, серотонина и гистамина в перитонеальной жидкости, брыжейке матки и периферической крови неравнозначно в течение периодов эстрального цикла. Динамика изменений концентрации биоаминов в исследованных элементах носит фазовый сопряженный во времени характер. Эта закономерность может отражать и определять интеграцию экстра- и интраорганных структур матки, обеспечивающих ее биоаминовый обмен, при переходах организма на новые уровни гомеостаза, соответствующие периодам половой цикличности.
СПИСОК ЛИТЕРАТУРЫ:
1. Абрамченко В.В., Капленко О.В. Адренергические средства в акушерской практике. – СПб. ТОО ТК «Петрополис». 2000. 272 с.
2. Бурлев В.А., Лец Н.И. //Проблемы репродукции. 2001. №1. С.25.
3. Виноградов С.Ю., Диндяев С.В., Погорелов Ю.В. и др. //Гистогенез и регенерация тканей. С.-Пт.: ВМА, 1995. С. 62.
4. Виноградов С.Ю., Погорелов Ю.В. //Вестник Ивановской мед. академии. 1997. Т.2. №3. С.6.
5. Гордон Д.С., Сергеева В.Е., Зеленова И.Г. Нейромедиаторы лимфоидных органов. Л.: Наука. 1982. 128 с.
6. Диндяев С.В. Клиническая морфология яичников. Иваново. 2005. 145с.
7. Диндяев С.В., Ткачева Е.А., Кротов А.С. //Актуальные проблемы биологии, медицины и экологии. Томск. 2004. Т. 3. № 1. С. 96.
8. Диндяев С.В., Виноградов С.Ю. //Бабухинские чтения в Орле 5-7 июня 2006 г. Москва, 2006. С. 47.
9. Диндяев С.В., Виноградов С.Ю. //Курский научно-практический вестник «Человек и его здоровье». 2006. №2. С.5.
10. Диндяев С.В., Стегнин М.В., Ткачева Е.А. и др. //Матер. науч. конфер. II Должановские морфологические чтения. Старый Оскол: ИПК «Кириллица», 2006. С. 74.
11. Лычкова А.Э. //Успехи физиол. наук. 2006. Т. 37. № 1. С. 50.
12. Csaba G., Kovacs P., Pallinger E. //Cell Biol. 2003. V. 27. № 4. P.387.
13. Csaba G., Kovacs P., Pallinger E. //Life Sci. 2006. V. 78. № 16. P.1871.
14. Martins E.Jr., Ferreira A.C., Skorupa A.L. et al. //J. Leukoc. Biol. 2004. V. 75. №6. P. 1116.
15. Metcalfe D.D., Baram D., Mekori Y.A. Mast cells //Physiol. Rev. 1997. V. 77. № 4. P. 1033.
16. Nakano K., Takamatsu S. //Nippon Yakurigaku Zasshi. 2001. V. 118. № 1. P. 15.
17. Rudd M.L., Nicolas A.N., Brown B.L. et al. //J. Neuroimmunol. 2005. V.159. № 1-2. P. 113.
18. Swindle E.J., Hunt J.A., Coleman J.W. //J. Immunol. 2002. V. 169. № 10. P. 5866.
Библиографическая ссылка
Диндяев С.В. ВНЕОРГАННЫЕ СТРУКТУРЫ В СИСТЕМЕ РЕГУЛЯЦИИ БИОАМИНОВОГО ОБМЕНА МАТКИ // Фундаментальные исследования. 2007. № 10. С. 15-19;URL: https://fundamental-research.ru/ru/article/view?id=4474 (дата обращения: 17.05.2026).