



До сих пор большинство работ по моделированию динамики пространственно распределённых биологических сообществ [3–5] основываются на уравнениях «реакция – диффузия», требующих, чтобы особи перемещались по пространству случайно и бесцельно, чего не наблюдается в живой природе. Принцип эволюционной оптимальности [1, 2] позволяет создавать модели, свободные от этого противоречия: особи перемещаются целенаправленно, максимизируя средний по пространству коэффициент размножения (КР). При целенаправленных миграциях большое значение имеет информация, которой располагают особи, принимая решение о перемещении. Рассматривают следующие модели информированности особей:
– глобальной информированности – особи обладают полной информацией о состоянии окружающей среды и численностях субпопуляций в обеих стациях, а также о цене перемещения (т.е. затраты на перемещение из стации в стацию, выраженные в долях КР);
– полной неинформированности;
– локальной информированности, в которой особи располагают частичной информацией, необходимой для принятия решения о миграции.
В настоящей работе будет рассмотрена модель с глобальной информированностью особей.
Модель оптимизационной миграции глобально информированных особей
Рассмотрим модель, описывающую пространственно распределенное двухвидовое сообщество, особи которого глобально информированы. Будем считать, что сообщество занимает две стации. Ситуацией миграции будет перемещение особей (хотя бы одного вида) из стации в стацию; все остальные перемещения внутри стаций миграцией являться не будут. Динамика каждой из популяций в каждой стации описывается разностным аналогом уравнения Лотки – Вольтерры:
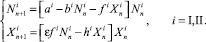 (1)
(1)
где Xn – численность хищников в момент времени n; Nn – численность жертв в момент времени n; a – характеризует плодовитость популяции жертв; b – характеризует площадь, которая необходима особи жертвы для нормального воспроизводства; h – аналогично b, но для хищников; f – успешность поиска и атаки хищниками жертв, а также успешность пользования жертвами убежищами либо защиты; ε – параметр, характеризующий эффективность превращения биомассы жертв в биомассу хищников (0 < ε < 1).
Вероятность перемещения без ущерба для дальнейшего размножения жертв (хищников) – цену перемещения – обозначим p (соответственно q). Для каждой стации определим коэффициенты размножения:
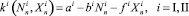 для жертв; (2)
для жертв; (2)
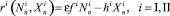 для хищников. (3)
для хищников. (3)
Особи начинают мигрировать, если условия существования в противоположной стации лучше, чем в текущей стации. Особи перемещаются таким образом, чтобы максимизировать (средний по пространству) коэффициент размножения (КР). Тогда условие миграции жертв в случае I → II описывается неравенством
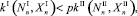 (4)
(4)
аналогично записывается условие для миграции II → I.
Условие начала миграции хищников в случае I → II описывается неравенством
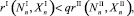 (5)
(5)
аналогично записывается условие для миграции I → II.
Величина миграционного потока выбирается таким образом, чтобы условия (4)–(5) превратились в равенство. Для хищников в случае I → II
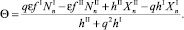 (6)
(6)
Величина миграционного потока для жертв (Δ) в случае I → II определяется как:
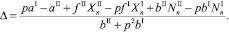 (7)
(7)
Если на каком-то шаге времени n ни одно из неравенств (4)–(5) не выполняется, то миграции на данном шаге времени не происходит.
Модель работает следующим образом: на каждом шаге времени t определяются величина и направление миграционных потоков Θ и Δ. Происходит перераспределение особей между стациями. Затем происходит размножение в силу системы уравнений (1) с теми численностями, которые складываются в результате перераспределения особей.
Условия существования стационарной динамики
Пусть оба вида не мигрируют, и значения всех численностей не меняется на каждом шаге времени. То есть, начиная с некоторого i > k, значения численностей на предыдущем шаге совпадают со значениями численностей на текущем шаге и равны NI, XI, NII, XII:
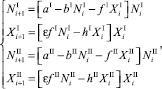
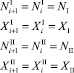 ∀i > k. (8)
∀i > k. (8)
Поскольку особи обеих стаций не взаимодействуют друг с другом, то можно рассматривать только одну из них. Тогда существуют такие стационарные значения NI, XI, что, начиная с некоторого i > k:
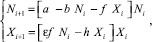
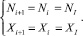 (9)
(9)
Рассмотрим все возможные стационарные случаи.
1. NI = 0, XI = 0. Этот случай вырожденный, поскольку если NI = 0, тогда ввиду отсутствия кормовой базы хищники вымирают и XI = 0.
2. NI ≠ 0, XI = 0. Имеем следующую стационарную численность жертв при вымирании хищников:
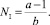 (10)
(10)
3. NI ≠ 0, XI ≠ 0. Тогда КР на каждом шаге времени, начиная с некоторого k > 0, равны единице:
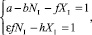 (11)
(11)
откуда можно получить значения стационарных численностей:
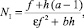
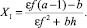 (12)
(12)
По условию a ≥ 1. Тогда для выживания хищников необходимо выполнение условия
εf(a – 1) > b. (13)
Утверждение: из системы (9) следует, что если один из видов имеет ненулевую стационарную динамику, то и второй вид также имеет стационарную динамику численности.
Доказательство: Пусть динамика численности жертв имеет стационарный характер. Тогда из уравнения для численности жертв следует, что $NI > 0 и некоторое k > 0, что ∀i > k NI = Ni и выполняется равенство
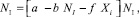 ∀i > k. (14)
∀i > k. (14)
Поскольку NI положительная константа, сократив (14) на NI и выразив Xi, получим выражение
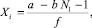 ∀i > k. (15)
∀i > k. (15)
Выражение в правой части постоянно ∀i > k, тогда Xi = XI тоже постоянная величина ∀i > k. Напротив, предположим, что численность хищников имеет ненулевую стационарную динамику. Тогда, из уравнения для численности хищников (9) следует, что $XI > 0 и некоторое k > 0, что ∀i > k XI = Xi и выполняется равенство:
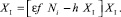 (16)
(16)
Так как XI положительная константа, сократив (16) на XI и выразив Ni, получим следующее выражение:
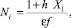 ∀i > k. (17)
∀i > k. (17)
Выражение в правой части постоянно ∀i > k, тогда Ni = NI также постоянная величина ∀i > k. Утверждение доказано.
Отметим, что случай вымирания одного из видов является вырожденным: если популяция жертв имеет нулевую стационарную динамику, то хищники, ввиду отсутствия ресурсов также имеют нулевую стационарную динамику; если же произошло вымирание популяции хищников, то динамика жертв может иметь любой характер.
Пусть теперь мигрирует один из видов. Предположим что динамика численности жертв и хищников стационарна, то есть значения численностей на предыдущем шаге совпадают со значениями численностей на текущем шаге, положительны и равны NI, XI, NII, XII. Пусть хищники не мигрируют, а жертвы мигрируют, например, из первой стации во вторую. Тогда система уравнений (1) примет следующий вид:
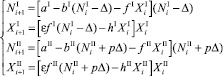
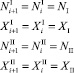 ∀i > k. (18)
∀i > k. (18)
Так как жертвы мигрируют из первой стации во вторую, то в соответствии с уравнением (2):
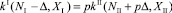 . (19)
. (19)
Это выражение можно переписать и обозначить через l:
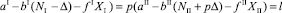 . (20)
. (20)
Подставив l в уравнения численности жертв системы (18), получим (заметим, что p > 0, иначе миграция отсутствует)
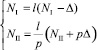 . (21)
. (21)
Поделив первое уравнение на второе, и сократив l (l > 0 всегда, поскольку значения стационарных численностей ненулевое) получим выражение
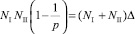 , (22)
, (22)
в левой части которого неположительное выражение, поскольку 0 < p ≤ 1. Пришли к противоречию, поскольку численности и величина миграционного потока по предположению положительные. Аналогично доказывается отсутствие миграции, если предположить миграцию хищников.
Одновременная миграция хищников и жертв
Пусть численность каждого вида в стациях на каждом шаге времени не меняется и положительна. Хищники мигрируют, жертвы тоже. Так как численность хищников и жертв не меняется, то несложно доказать, что каждый вид мигрирует всегда в одном направлении.
Пусть хищники и жертвы мигрируют из первой стации во вторую. Поскольку мы рассматриваем случай стационарных значений численностей, то, начиная с некоторого i > k, значения численностей на предыдущем шаге совпадают со значениями численностей на текущем шаге и равны NI, XI, NII, XII:
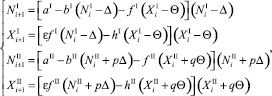
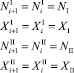 ∀i > k. (23)
∀i > k. (23)
Поскольку хищники мигрируют из первой стации во вторую, то в соответствии с (2) имеем
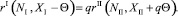 (24)
(24)
Перепишем это уравнение в следующем виде и обозначим выражения через m:
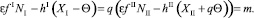 (25)
(25)
Подставив m в уравнения (23), получим (при условии q > 0)
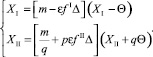 (26)
(26)
Выразив m и приравняв уравнения, получим
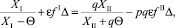
откуда
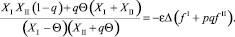 (27)
(27)
Поскольку правая часть (28) не положительна, а левая всегда положительна (ввиду положительности всех численностей), то Δ = 0. Если Δ = 0, то (28) примет вид
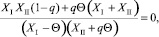
откуда
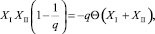 (28)
(28)
что имеет смысл только при Θ = 0.
Мы имеем Δ = 0 и Θ = 0, что означает отсутствие миграции. Таким образом, для рассматриваемой модели мы доказали следующую теорему. Теорема о стационарной динамике: если динамика численности особей стационарна, то для такого режима миграции не существует.
Устойчивость стационарной динамики
Поскольку модель является многопараметрической, поиск самих предельных режимов представляет собой сложную задачу. Мы ограничимся рассмотрением устойчивости стационарного режима в малом приближении.
Поскольку мы предполагаем, что в двух стациях динамика стационарна, то условия миграции (4)–(5) выполняться не будут, поскольку КР равен единице. Для исследования устойчивости системы применим линейный анализ. Положим
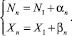 (29)
(29)
Линеаризуем уравнения (1), разлагая функцию по степеням αn, βn и отбрасывая члены второго порядка и выше. Обозначим μn = (αn, βn)T, тогда получим
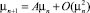 , (30)
, (30)
где A – матрица коэффициентов при членах первого порядка; 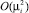 – матрица членов порядка 2 и больше. Подставим (30) в (29) и приведем к матричному виду:
– матрица членов порядка 2 и больше. Подставим (30) в (29) и приведем к матричному виду:
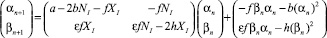 . (31)
. (31)
Так как КР равны единице, матрица A имеет вид
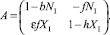
Откуда корни характеристического уравнения A – λE = 0 равны
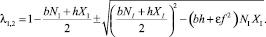 (32)
(32)
Пусть λi, 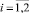 – собственные числа матрицы A. Как известно, система асимптотически устойчива тогда и только тогда, когда модули всех корней (полюсов) характеристического уравнения системы меньше 1, т.е.
– собственные числа матрицы A. Как известно, система асимптотически устойчива тогда и только тогда, когда модули всех корней (полюсов) характеристического уравнения системы меньше 1, т.е. 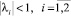 . Тогда условие асимптотической устойчивости системы таково:
. Тогда условие асимптотической устойчивости системы таково:
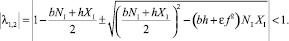 (33)
(33)
Таким образом, если параметры системы удовлетворяют выражению (34), то стационарная динамика является устойчивой.
Заключение
Мы показали, что для модели глобально информированных особей двухвидового пространственно распределенного сообщества для каждого набора параметров существует стационарная динамика. Причем стационарная динамика возможна только в случае отсутствия миграции особей обоих видов. Даны оценки устойчивости стационарной динамики системы относительно изменения начальных данных.
Рецензенты:
Заворуев В.В., д.б.н., ведущий научный сотрудник Института вычислительного моделирования СО РАН, г. Красноярск;
Лейнертас Е.К., д.ф.-м.н., профессор кафедры ТФКП ИМиФИ, СФУ, г. Красноярск.
Работа поступила в редакцию 06.11.2014.
Библиографическая ссылка
Сенашова М.Ю., Садовский М.Г. СУЩЕСТВОВАНИЕ И УСТОЙЧИВОСТЬ СТАЦИОНАРНОЙ ДИНАМИКИ ПОПУЛЯЦИИ ДВУХВИДОВОГО СООБЩЕСТВА ПРИ ОПТИМИЗАЦИОННОЙ МИГРАЦИИ // Фундаментальные исследования. 2014. № 11-7. С. 1546-1551;URL: https://fundamental-research.ru/ru/article/view?id=35804 (дата обращения: 20.03.2026).