



Наибольшим воздействиям и изменениям организм подвержен на ранних стадиях эмбриогенеза. Отсюда понятен огромный интерес, который проявляется к эмбриологическим исследованиям. Происхождение различных форм пороков органов репродуктивной системы зависит от того, на каком этапе эмбриогенеза оказал действие тератогенный фактор или реализовалась наследственная патология [2,5,6,10]. В связи с этим необходимо глубоко понимать не только ход самого процесса эмбрионального развития тканей и органов, но и роль базисных закономерностей гистогенеза и их органоспецифические особенности [3,7,9]. Анализ данных литературы показывает, что вопросы гистогенеза тканей влагалища, образующих стенку органа, а особенно мышечных, изучены недостаточно [1,4, 8]. Полученные данные нуждаются в дальнейшем накоплении фактического материала.
Цель работы – изучить ход эмбрионального гистогенеза мышечных тканей влагалища.
Материалы и методы исследования
В работе использован материал от плодов крыс женского пола с 14,5 по 21-е сутки пренатального развития в соответствии с «Правилами проведения работ с использованием экспериментальных животных». Для проведения светового исследования использовали фиксацию материала в 10 % нейтральном формалине на фосфатном буфере (рН-7,4), заливку в парафин. Для электронной микроскопии использовали префиксацию в 2,5 % глутаральдегиде на 0,2 М какодилатном буфере (рН 7,4), фиксацию в 1 % ОsО4 и заливали в аралдит. Для обеспечения прицельного электронно-микроскопического анализа получали серийные полутонкие срезы толщиной 1-2 мкм, которые окрашивали 1 % раствором метиленового синего. Прицельные ультратонкие срезы толщиной 200-500 нм просматривали в электронном микроскопе Hitachi-HU-12.
Результаты исследования и их обсуждение
Во время органной дифференцировки (14,5-15,5 сутки развития) у плодов крыс, матка и влагалище приобретают характерные черты для каждого органа. Многослойный плоский эпителий является уникальным для влагалища, происходящего из Мюллерова протока, нечувствительного к действию АМГ. Эпителий спонтанно подвергается трансформации из напоминающего маточный, цилиндрического, в многослойный плоский. У крыс период чувствительности к АМГ («окно» чувствительности) приходится на 14-15 сутки внутриутробного развития. В развивающихся органах репродуктивной системы, как и во многих других (кишечник, мочевой пузырь и др.), мезенхимальные клетки в тесной близости к эпителию дифференцируются в фибробласты, чтобы образовать слизистую оболочку, тогда как наиболее отдаленные дифференцируются в гладкие миоциты, формируя мышечную оболочку.
В области формирования мышечной оболочки влагалища клетки располагаются как одиночно, так и небольшими скоплениями. Соседние клетки часто образуют протяженные простые контакты, интердигитации, а иногда и щелевидные контакты. Большая часть клеток имеет форму близкую к округлой, некоторые несколько вытянуты. Они несколько отличаются по ядерно-цитоплазматическому соотношению, но большинство характеризуется его высоким соотношением. В таких клетках ядра чаще округлые, с небольшими инвагинациями оболочки и высоким содержанием эухроматина. Ядрышки наблюдаются не часто и обычно не крупные. В цитоплазме встречаются немногочисленные митохондрии, единичные короткие канальца гранулярной ЭПС, много свободных рибосом и гранул гликогена. Иногда наблюдаются единичные диктиосомы комплекса Гольджи. По своей ультраструктурной организации они соответствуют премиобластам (рис. 1).
О запуске процессов морфологической дифференцировки свидетельствует более быстрое накопление органелл общего значения. Кроме большого количества свободных рибосом, в этих клетках становится больше канальцев гранулярной ЭПС, появляются диктиосомы комплекса Гольджи. Крупные рыхлые ядрышки также подтверждают интенсификацию синтетических процессов. Морфология подавляющего большинства клеток не позволяет достоверно судить об их тканеспецифичности.
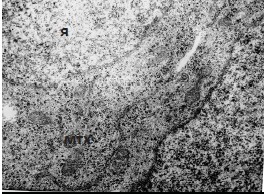
Рис. 1. Ультраструктурная организация премиобластов, 15-е сутки эмбрионального развития. Я – ядро, мх – митохондрии. Ув. 12000.
Однако их компактизация с установлением простых контактов и десмосом позволяет рассматривать эти клетки как предшественники мышечных элементов. На 16-е сутки эмбрионального развития крыс при электронно-микроскопическом исследовании удается обнаружить появление миофиламентов. Это дает возможность идентифицировать часть клеток как миобласты. Они характеризуются снижением ядерно-цитоплазменного соотношения за счет увеличения объема цитоплазмы. Миофиламенты расположены небольшими скоплениями, обычно ближе к периферии клетки, вдоль ее длинной оси, между ними располагается большое количество рибосом и полисом. Таким образом, проявления дивергентной дифференцировки мезенхимных клеток дистальных отделов парамезонефральных протоков в элементы гладкой мышечной ткани отмечаются на 15-16 сутки, становясь очевидными на 17 сутки эмбрионального развития.
На 17-е сутки эмбрионального развития влагалище представляет собой трубку, растущую к преддверию (рис. 2).
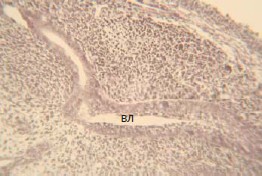
Рис. 2. Фрагмент стенки влагалища, 17 сутки эмбрионального развития крысы. Окраска по Ван-Гизон. Об. 20, ок. 10.
В просвете органа имеются слущенные клетки, многослойный плоский эпителий имеет признаки вертикальной анизоморфии. В подлежащей ткани мезенхима активно дифференцируется в соединительнотканные и мышечные элементы.
К концу 17 суток миобласты вытягиваются, в них наблюдается накопление органелл общего значения, концентрирующихся ближе к полюсам ядра. Начинается формирование кластеров из двух-трех клеток с образованием протяженных простых контактов и десмосом между ними. На 18-е сутки пренатального развития происходит дальнейший рост органа. В области развивающейся мышечной оболочки клетки компактизируются, можно достаточно четко определить ее границы. Миобласты приобретают вытянутую форму и начинают объединяться в группы. Соседние клетки в компартменте формируют, наряду с простыми контактами, нексусы и десмосомы. В популяции лейомиоцитарного ряда усиливается гетероморфность и гетерохронность морфологической дифференцировки. Это подтверждается обнаружением клеток на разном этапе процесса специфической дифференцировки в первую очередь, а также разной синтетической активностью клеток. Так, ядра миобластов отличаются разным количеством гетерохроматина и размером ядрышек. Хотя в среднем можно говорить о высокой синтетической активности клеток этой популяции. Как следствие разной интенсивности синтетических процессов клетки содержат и разное количество миофиламентов. Их уровень развития соответствует малодифференцированным лейомиоцитам. Большая плотность расположения филаментов создает высокую электронную плотность цитоплазмы и такие клетки выглядят более темными. При этом, накопив больше сократительных структур, они содержат несколько меньше органелл общего значения, в первую очередь ЭПС и комплекса Гольджи.
Ближе к периоду рождения, с увеличением возраста плодов, по мере увеличения количества миофиламентов в клетках, они располагаются в цитоплазме все более равномерно. В участках начала их синтеза, они располагаются в виде небольших пучков, обычно на периферии клетки. С увеличением количества сократительных структур, они занимают не только периферическую часть клетки, но и области цитоплазмы, расположенные ближе к ядру. Лейомиоциты с подобной ультраструктурной организацией можно отнести к дифференцирующимся клеткам.
Формирование мышечной оболочки к 19-м суткам эмбрионального развития, наряду с дифференцировкой клеток, сопровождается и элиминацией части клеток. Среди нормально развивающихся клеток сравнительно часто отмечаются клетки в состоянии апоптоза (рис.3).

Рис. 3. Ультраструктурная организация мышечной оболочки с большим количеством апоптотических клеток (ап), 19-е сутки эмбрионального развития. Ув. 3600.
Вероятно, это объясняется конкуренцией среди миобластов за факторы, влияющие на их рост и дифференцировку. По нашему мнению, источником подобных факторов на этом этапе гистогенеза могут являться нервные элементы.
К концу эмбриогенеза становится возможным наблюдать образование симпластических структур поперечно-полосатого компонента мышечной оболочки. Дистальнее от эпителия, ближе к наружи, в нижней трети влагалища, определяются скопления клеток, напоминающих своим строением премиобласты. Они имеют ядро округлой или овальной формы, дисперсно расположенный хроматин. Цитоплазма слабо развита, содержит единичные неразвитые митохондрии, небольшое количество рибосом, полисом, небольшие цистерны ЭПС, комплекс Гольджи. Дифференцируясь, эти клетки становятся миобластами. Их размеры увеличиваются, развивается цитоплазма и органеллы общего значения. Так же, как и дифференцировка гладкомышечных элементов, дифференцировка мышечных волокон характеризуется гетерохронностью. Слияние клеток, образование мышечных трубочек происходит, по-видимому, за счет постепенного разрушения небольших участков плазмолемм и объединения цитоплазмы. Вероятно, есть и другой механизм слияния миобластов. Некоторые клетки сближаются, не образуя контактов, и между ними остается щелевидное пространство. Клетки в нескольких участках сразу образуют небольшие выпячивания, комплементарные подобным же инвагинациям плазмолеммы соседней клетки. В этих участках и начинается слияние мембран клеток. Практически сразу после этого начинается активный миофибриллогенез. «Пионерские» миофибриллы первоначально располагаются в перинуклеарной области или в глубине цитоплазмы (рис. 4). Дальнейшее накопление миофиламентов приводит к удлинению и утолщению миофибрилл. В них появляются плохо выраженные Z-полоски.
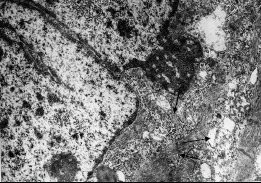
Рис. 4. Ультраструктурная организация первых миосимпластов мышечной оболочки влагалища, 20-е сутки эмбрионального развития. ↑ – миофибриллы. Ув. 10000.
На 21 сутки пренатального гистогенеза темп специфической дифференцировки нарастает и в стенке влагалища можно обнаружить элементы разной степени дифференцировки гладкой и поперечно-полосатой мышечной тканей. Обнаруживается послойная ориентация гладкомышечных клеток.
Таким образом, тканевой состав мышечного компонента стенки влагалища имеет различные (гетерогенные) эмбриональные источники развития. Обнаружение клеток мезенхимной природы в области закладки мышечной оболочки влагалища показывает их неоднородность: можно различить «темные» и «светлые» мезенхимные клетки, окружающие эпителий. По характеру удаленности от эпителия, выстилающего влагалище, можно предположить, что «темные» мезенхимные клетки можно рассматривать как премиобласты, исходя из их последующей клеточной дифференцировки. «Светлые» мезенхимные клетки, локализуясь субэпителиально, дифференцируются, в дальнейшем, в фибробласты. Для них характерно отсутствие каких-либо контактных взаимодействий и высокий внутриклеточный объем цистерн ЭПС. В ходе дифференцировки лейомиоцитов происходит не только усложнение ультраструктурной организации и формирование различных фенотипов лейомиоцитов, но и усложнение уровня межклеточных контактов.
Источником развития исчерченной мышечной ткани служат миобласты, которые скорее всего мигрировали из зачатков мышц промежности. Гетерохронность развития мышечных элементов в процессе специфической дифференцировки наиболее отчетливо проявляется в период слияния миобластов (даже при еще не полном объединении), когда некоторые клетки уже содержат толстые длинные миофибриллы, а часть миобластов лишена сократительных структур. В процессе специфической дифференцировки для исчерченной мышечной ткани влагалища характерна последовательная смена стадий развития: премиобласты, миобласты, мышечные трубочки и миосимпласты. На миотомное происхождение данного вида ткани указывает также наличие миосателлитоцитов в мышечных волокнах.
Рецензенты:
Суворова Г.Н., д.м.н., профессор, заведующая кафедрой анатомии человека, ГБОУ ВПО «Самарский государственный медицинский университет» Минздрава России, г. Самара;
Колсанов А.В., д.м.н., профессор, заведующий кафедрой оперативной хирургии и клинической анатомии человека с курсом инновационных технологий ГБОУ ВПО «Самарский государственный медицинский университет» Минздрава России, г. Самара.
Работа поступила в редакцию 24.06.2014.
Библиографическая ссылка
Шурыгина О.В., Ямщиков Н.В., Абрамов В.Н., Балашов В.П. ЭМБРИОНАЛЬНОЕ РАЗВИТИЕ МЫШЕЧНЫХ ТКАНЕЙ СТЕНКИ ВЛАГАЛИЩА КРЫС // Фундаментальные исследования. 2014. № 7-4. С. 812-816;URL: https://fundamental-research.ru/ru/article/view?id=34988 (дата обращения: 01.07.2025).