



На сегодняшний день трудно недооценить роль пчелы медоносной как для биоценозов, так и для человека. Пчелы обитают на значительных территориях с разным климатом, и успешно адаптируются к его особенностям. Для человека пчела полезна не только как источник ценных продуктов пчеловодства, но и как весьма интересный объект с точки зрения биологической науки. Обитая в пределах самых разнообразных ландшафтов с сильно отличающимися климатическими характеристиками пчелы в ходе естественного эволюционного процесса, под действием факторов среды приобрели определенные приспособительные механизмы, позволяющие им использовать ресурсы наиболее эффективно.
В результате сформировались расы пчелы медоносной, отличающиеся годичными биоритмами, этологическими и морфологическими особенностями [2]. Поскольку медоносная пчела – одно из самых изученных на сегодняшний день беспозвоночных животных, эти факты достаточно подробно освещены в литературных источниках [2, 3, 7, 9]. Несомненно, эти признаки закреплены генетически и затрагивают как внешнее строение, так и внутреннее, на уровне органов, тканей и клеток [6].
Для растительных сообществ огромное значение играют насекомые-опылители, в том числе пчелы, пчелиные семьи могут включать много тысяч особей и значительно влиять на жизнь местного биоценоза. Пермский край включает районы (напр. Уинский), которые являются местами обитания естественных популяций темной европейской лесной пчелы, т.е. среднерусской. Пчелам среднерусской расы, как известно, характерна наибольшая зимостойкость, которая определяет возможность их обитания на северных границах ареала, а также является чрезвычайно полезным для пчеловодческой отрасли качеством [2, 9].
Для Пермского края характерен длительный безоблетный период с продолжительным воздействием низких температур и весенние возвратные заморозки, как правило, отрицательно сказывающиеся на силе пчелиных семей. Более теплолюбивые расы медоносной пчелы, например карпатская, завозимые пчеловодами имеют положительные этологические характеристики, такие как меньшая злобливость, активность при низком уровне взятка, но уступают по зимостойкости аборигенной среднерусской расе [2, 8].
Расовые группы медоносных пчел имеют достаточно много различий, установленных разными исследователями, принимаемых во внимание при организации современных пчеловодческих хозяйств и важных для понимания биологии пчел [5, 7, 8, 9].
Тема гистологических исследований весьма актуальна и позволяет дополнить общие представления о биологии перепончатокрылых насекомых и в частности пчелы медоносной (Apis mellifera L.)
Интереснейшей отличительной особенностью медоносных пчел от большинства других перепончатокрылых является их способ зимовки. В течение всего безоблетного периода насекомые находятся в активном состоянии, не впадая в настоящий анабиоз, поддерживая положительную температуру внутри клуба за счет сокращений летательной мускулатуры. Такой способ зимовки является результатом длительной эволюции и подразумевает наличие комплексных приспособительных механизмов.
Успех зимовки пчелиной семьи зависит от многих внутренних и внешних факторов, например, количество и качество запасенных в гнезде кормовых ресурсов, а также состояния самих рабочих пчел и уровень их физиологической изношенности. В течение года происходит плавная последовательная смена поколений имаго рабочих пчел. Онтогенез каждого поколения происходит при различных условиях, несмотря на относительно стабильные условия в гнезде, может сильно отличаться состав кормов, количество расплода, физиологическая нагрузка на летных и ульевых пчел (молодые имаго).
Летние поколения рабочих особей в среднем проживают 35–40 дней, так как изнашиваются во время медосбора, осенняя генерация уходящая в зимовку проживает в среднем до 7–8 месяцев, так как практически не участвует в фуражировочной деятельности.
В течение всего безоблетного периода пчелы потребляют запасенные в гнезде корма для поддержания необходимой температуры, что особенно важно в начале весенних месяцев, когда начинается закладка весенней генерации рабочих особей. Эффективное использование кормов обеспечивается активными обменными процессами в средней кишке (ventriculus). Данная работа посвящена исследованию годовых изменений структуры вентрикулярного эпителия у пчел среднерусской расы.
Материалы и методы исследований
Для исследований использовались имаго рабочих особей пчелы медоносной из племенных пчелосемей пасеки компании «Парасоль», пасеки на территории села Бершеть Пермского края, а также исследовательской пасеки ПГПУ.
Забор проб производился с интервалом в среднем около 5–6 дней, т.е. 1–2 раза в неделю, на протяжении 2007–2013 гг. Фиксация и заливка образцов в парафин производилась автоматически на гистопроцессоре, в соответствии со стандартной методикой, адаптированной для данного вида материала [1]. Для изготовления срезов толщиной в 10 мкм использовался санный микротом МС-2, а также ротационный «Ротмик-1». В качестве монтирующей среды применялся пихтовый бальзам, а также синтетическая среда «Glasseal». Окраска срезов средней кишки производилась по стандартной методике гематоксилин – эозин [4]. Препараты были исследованы с помощью микроскопа «Микмед-6» и документировались камерой DCM -900. Микрометрические исследования и статистическая обработка данных производились на ПК c использованием Microsoft Excel 2003.
В силу энтодермального происхождения средняя кишка (ventriculus) не имеет хитиновой кутикулы, как передний и задний отделы кишечника, ее выстилка представлена однослойным столбчатым железистым эпителием. Клеточными элементами этой ткани обеспечивается две важнейшие для организма насекомого функции – синтез перитрофической мембраны и секреция пищеварительных ферментов, а также всасывание продуктов пищеварения. Оба этих процесса осуществляются сходными по строению клеточными элементами, без выраженной морфологической дифференциации лишь пространственно разобщенными по длине кишки [3, 5, 8]. Согласно данным многих исследователей эпителий средней кишки насекомых, в том числе и пчелы медоносной, имеет сложную ячеистую структуру. Стенка кишки образует многочисленные складки, несущие на вершинах железистые клетки, кроме того, в чашеобразных вдавливаниях, чередующихся с ними, располагаются крипты – центры регенерации клеток эпителия [2, 3, 11].
Эпителиальные клетки за свою жизнь проходят несколько условных стадий формирования, каждой из которых соответствует различный уровень секреторной активности.
Последняя стадия отличается образованием на апикальных концах клеток характерных колбовидных утолщений, пузырьков, наполненных секретом – апокринных головок.
Данные образования, отделяясь, попадают в просвет кишки, где лопаются, изливая пищеварительные ферменты, их количество на срезе отражает уровень секреторной активности на данном участке кишки [3, 5, 8].
При анализе изображения поперечного среза средней кишки производился подсчет упомянутых структур на 1/4 поперечного среза средней кишки, после чего данная величина соотносилась с единицей длины окружности среза (мм). Таким образом, физиологические колебания диаметра кишки не влияют на относительное количество апокринных головок. Кроме того, производились измерения диаметра кишки, высоты эпителиальных клеток в 3 участках среза с последующим усреднением (т.к. толщина эпителия в пределах среза может несколько отличаться). Упомянутые апикальные утолщения клеток хорошо окрашиваются гематоксилином, что позволяет производить их подсчет с высокой точностью [10].
При построении годовой картины изменений, происходящих в эпителиальном слое средней кишки, производилось объединение результатов измерений нескольких проб, в результате объем выборки, соответствующей одному месяцу, составлял от 10 до 15 имаго рабочих пчел.
Результаты исследования динамик измеряемых величин приводились и в более ранних исследованиях, но по отдельности изменения этих параметров достаточно сложно анализировать [5].
Впоследствии были введены комплексные параметры, учитывающие размеры элементов эпителия, и соотношение объема «секреторной трубки» кишки и количества концевых пузырьков, характеризующих активность секреции.
Далее приводится картина годовых изменений полного объема секреторной трубки, т.е. всего совокупного объема эпителиальной ткани средней кишки (Vs), а также секреторного коэффициента, характеризующего отношение объема эпителиальной ткани к количеству апокринных головок в тысячах (Sq).
Увеличение значения параметра Vs характеризует увеличение длины всего отдела в целом, либо рост толщины эпителия.
Необходимость такого параметра, как средняя длина кишки кажется необходимой, но кишка имеет физиологические складки, обусловленные строением стенки и работой мускулатуры, и очевидно, что длина отдела может меняться.
Результаты исследования и их обсуждение
В табл. 1 и 2 представлены годовые изменения исследуемых параметров, Показатели за каждый месяц сравнивались с предыдущим, январь – с декабрем.
Таблица 1
Изменения диаметра средней кишки и средней толщины вентрикулярного эпителия в течение года у имаго рабочих особей пчел среднерусской расы (P < 0,05, n = 10)
|
Месяц |
D – диаметр кишки (мм) |
h – толщина эпителия (мм) |
||||
|
X ± m |
d |
t |
X ± m |
d |
t |
|
|
Январь |
1,170 ± 0,05786 |
– |
– |
0,070 ± 0,00153 |
0,012 (16,6 %) |
2,82 |
|
Февраль |
0,936 ± 0,03102 |
–0,234 (20,0 %) |
3,57 |
0,072 ± 0,00235 |
– |
– |
|
Март |
1,070 ± 0,04330 |
0,134 (14,3 %) |
2,51 |
0,078 ± 0,00457 |
– |
– |
|
Апрель |
0,872 ± 0,02985 |
–0,197 (18,5 %) |
3,75 |
0,065 ± 0,00472 |
– |
– |
|
Май |
1,077 ± 0,05893 |
0,205 (23,5 %) |
3,10 |
0,046 ± 0,00136 |
–0,019 (28,8 %) |
3,82 |
|
Июнь |
1,009 ± 0,05454 |
– |
– |
0,079 ± 0,00840 |
0,033 (70,6 %) |
3,85 |
|
Июль |
0,877 ± 0,03073 |
-0,132 (13,1 %) |
2,11 |
0,044 ± 0,00133 |
–0,035 (44,8 %) |
4,17 |
|
Август |
0,699 ± 0,02942 |
-0,178 (20,3 %) |
4,19 |
0,071 ± 0,00430 |
0,027 (62,7 %) |
6,09 |
|
Сентябрь |
0,970 ± 0,02755 |
0,271 (38,7 %) |
6,72 |
0,059 ± 0,00258 |
–0,012 (17,6 %) |
2,49 |
|
Октябрь |
0,913 ± 0,05607 |
– |
– |
0,059 ± 0,00399 |
– |
– |
|
Ноябрь |
1,114 ± 0,04045 |
0,202 (22,1 %) |
2,92 |
0,064 ± 0,00461 |
– |
– |
|
Декабрь |
1,109 ± 0,04827 |
– |
– |
0,058 ± 0,00383 |
– |
– |
Таблица 2
Изменения общего объема вентрикулярного эпителия и его секреторного коэффициента в течение года у имаго рабочих особей пчел среднерусской расы (P < 0,05, n = 10)
|
Месяц |
Vs – Объем эпителия (мм3) |
Sq – Секреторный коэффициент (тыс. шт./мм3) |
||||
|
X ± m |
d |
t |
X ± m |
d |
t |
|
|
Январь |
2,387 ± 0,14840 |
0,600 (25,1 %) |
2,53 |
90,454 ± 7,87390 |
– |
– |
|
Февраль |
1,987 ± 0,10695 |
–0,400 (16,8 %) |
2,19 |
100,326 ± 5,76083 |
– |
– |
|
Март |
2,212 ± 0,17344 |
– |
– |
61,677 ± 6,83530 |
–38,649 (38,5 %) |
4,32 |
|
Апрель |
1,830 ± 0,10468 |
– |
– |
110,125 ± 6,96554 |
48,448 (78,6 %) |
4,96 |
|
Май |
2,118 ± 0,13326 |
– |
– |
124,546 ± 5,09187 |
– |
– |
|
Июнь |
3,350 ± 0,43688 |
1,232 (58,2 %) |
2,70 |
64,424 ± 10,23508 |
–60,122 (48,3 %) |
5,26 |
|
Июль |
1,071 ± 0,06350 |
–2,279 (68,0 %) |
5,16 |
106,884 ± 7,96111 |
42,460 (65,9 %) |
3,27 |
|
Август |
0,763 ± 0,04576 |
–0,307 (28,7 %) |
3,93 |
84,624 ± 4,45647 |
–22,260 (20,8 %) |
2,44 |
|
Сентябрь |
1,553 ± 0,08255 |
0,790 (103,5 %) |
8,37 |
85,400 ± 5,74819 |
– |
– |
|
Октябрь |
1,408 ± 0,07268 |
– |
– |
101,937 ± 4,93312 |
16,537 (19,4 %) |
2,18 |
|
Ноябрь |
1,884 ± 0,09158 |
0,477 (33,9 %) |
4,08 |
67,721 ± 6,37558 |
–34,216 (33,6 %) |
4,24 |
|
Декабрь |
1,787 ± 0,18523 |
– |
– |
83,981 ± 7,43027 |
– |
– |
Как видно из таблицы, повышение значения параметра приводит к положительной разности, понижение к отрицательной. Отличия, имеющие низкую статистическую достоверность, не приводятся. На рисунке представлены графики изменений исследуемых параметров.
Из-за сильно отличающихся пределов изменения параметров их графики представлены на разных рисунках, с единой разметкой по оси абсцисс, что позволяет соотносить точки разных кривых. Из таблицы видно, что наибольший диаметр средней кишки соответствует январю (1,170 ± 0,05786), значение толщины эпителия в этой точке слабо отличается от ближайших, объем эпителия (Vs) показывает слабый пик (2,387 ± 0,14840), такие значения могут свидетельствовать об укорочении среднего отдела кишечника, некоторой его компактизации (рисунок, б).
а 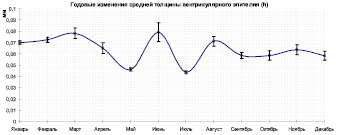 б
б 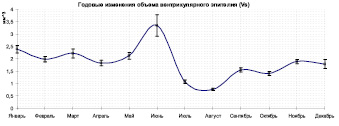
в 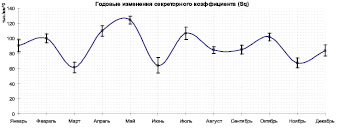
Годовые изменения средней толщины вентрикулярного эпителия (а), объема вентрикулярного эпителия (б) и секреторного коэффициента (в)
Данные об изменении диаметра кишки приводятся в обзорных целях, ventriculus у насекомых играет роль желудка, его форма может физиологически изменяться в зависимости от разных причин, таких как тип кормов, скорость пищеварения, уровень метаболизма насекомого. Некоторыми авторами описывается движение физиологических складок вдоль данного отдела кишечника на свежеотпрепарированных образцах, следовательно, форма средней кишки нестабильна, и ее метрические характеристики по всей видимости не могут дать полного представления о функциональной активности [3, 7, 8]. Изменение структуры клеточных элементов эпителия – более длительный и сложный процесс, секреторная активность этой ткани, по всей видимости коррелирует с основными онтогенетическими изменениями в организме насекомого, которые сопряжены со сменой фаз жизненного цикла пчелиной семьи.
Так, минимумы на графике значений секреторного коэффициента соответствуют таким месяцам, как март, июнь и ноябрь (61,677 ± 6,83530; 64,424 ± 10,23508; 67,721 ± 6,37558 соответственно) и по всей видимости могут быть связаны с присутствием большого количества молодых особей, питающихся кормами, не требующими обильной секреции в средней кишке. Как видно из графика толщины эпителия (h), т.е. высоты столбчатых железистых клеток, 2 из 3 пиков секреторного коэффициента соответствуют спаду значений этой величины – в мае (0,046 ± 0,00136 мм) и в июле (0,044 ± 0,00133 мм), из соотношения этих данных следует, что секреторный коэффициент увеличивается главным образом за счет уменьшения объема эпителиальной ткани.
Общий объем секреторной ткани (Vs) по всей вероятности связан как с уровнем секреции, так и всасывания, и может иллюстрировать также степень физиологического износа рабочих особей, что подтверждается полученными данными – например, в мае – июне наблюдается достоверное возрастание параметра Vs (3,350 ± 0,43688), что соответствует выходу значительной части молодых имаго весеннее-летней генерации. Затем следует спад (0,763 ± 0,04576), в июле-августе объем эпителия принимает наименьшее значение (рабочие имаго среднерусских пчел быстро изнашиваются во время активного медосбора).
При анализе динамик следует иметь в виду, что значительная часть жизненного цикла пчелиной семьи (с сентября-октября по апрель-май) протекает без смены генераций рабочих особей и во многом определяется онтогенезом осенней генерации, идущей в зимовку.
Выводы
1. На графике объема эпителия (Vs) заметно, что в течение периода с ноября по апрель наблюдается медленная деградация эпителиального слоя, по всей видимости связанная с медленным физиологическим износом рабочих особей в течение зимовки, и в итоге началом весенней фуражировочной деятельности (май).
2. В мае – июне пик значений этого параметра (Vs), вероятно, обусловлен выходом весенней генерации и затем ее достаточно быстрым физиологическим износом: во время медосбора одновременно гибнет значительная часть имаго, вышедших из зимовки.
3. Наибольшие значения секреторного коэффициента (Sq) в большинстве соответствуют выходу молодых имаго следующей генерации, им характерны более тонкие стенки эпителиальной трубки, что может быть также связано с кормовыми особенностями.
В результате данного исследования были выявлены достоверные изменения в строении средней кишки и секреторной активности вентрикулярного эпителия у рабочих имаго пчел среднерусской расы в течение года.
Наиболее динамично характеристики среднего отдела кишечника меняются в периоды, соответствующие ключевым изменениям в жизненном цикле пчелиной семьи.
Рецензенты:
Золотухин И.А., д.т.н., доцент, профессор кафедры ботаники ПГГПУ, г. Пермь;
Петрухин Ю.А., д.б.н., доцент, профессор кафедры ботаники ПГГПУ, г. Пермь.
Работа поступила в редакцию 06.03.2014.
Библиографическая ссылка
Лаврский А.Ю. ГОДОВАЯ ДИНАМИКА АКТИВНОСТИ ВЕНТРИКУЛЯРНОГО ЭПИТЕЛИЯ У ИМАГО РАБОЧИХ ОСОБЕЙ МЕДОНОСНЫХ ПЧЕЛ СРЕДНЕРУССКОЙ РАСЫ (APIS MELLIFERA MELLIFERA L.) // Фундаментальные исследования. 2014. № 3-4. С. 750-754;URL: https://fundamental-research.ru/ru/article/view?id=33748 (дата обращения: 18.07.2026).