


Scientific journal
Fundamental research
ISSN 1812-7339
"Перечень" ВАК
ИФ РИНЦ = 1,798
β-SITOSTEROL-TRANSFORMING ACTIVITY OF NATIVE AND MUTANT RHODOCOCCUS STRAINS

Каталитические свойства микробных ферментов позволяют эффективно трансформировать труднодоступные для химической модификации области молекулы природных стеролов. Микробиологическая трансформация данных соединений в андрост-4-ен-3,17-дион и андроста-1,4-диен-3,17-дион – ключевые интермедиаты в синтезе лекарственных гормональных препаратов является традиционной технологией, применяемой в промышленном производстве стероидов [6]. В исследованиях по микробиологической трансформации стеролов особое внимание уделяется представителям, выделяемым из отходов деревообрабатывающей промышленности (β-ситостеролу, в частности). Преимущество использования β-ситостерола и его аналогов заключается в тотальном распространении их в природе, доступности и относительно низкой стоимости исходного сырья [6]. Область исследования каталитических возможностей микроорганизмов в отношении данных соединений постоянно пополняется новыми данными, исследуется трансформирующая способность ранее мало используемых бактериальных штаммов и их мутантных клонов, проводится оптимизация биотрансформационного процесса в условиях добавления высоких концентраций стеролов и т. п. Ранее нами [1, 2, 4] с использованием нативных штаммов родококков, поддерживаемых в Региональной профилированной коллекции алканотрофных микроорганизмов (официальный акроним коллекции ИЭГМ, номер во Всемирной федерации коллекций культур 768) [7], исследован процесс биотрансформации β-ситостерола в концентрациях 0,5–2,0 г/л. Определены основные метаболические пути его модификации, при этом в качестве продуктов обнаружены биологически активные соединения стигмаст-4-ен-3-он, андрост-4-ен-3,17-дион, андроста-1,4-диен-3,17-дион и ацетат β-ситостерола.
Цель настоящей работы – исследование трансформирующей активности нативных и мутантных штаммов родококков в отношении β-ситостерола в зависимости от его исходной (0,5-10 г/л) концентрации.
Материалы и методы исследования
В работе использовали чистые культуры родококков из Региональной профилированной коллекции алканотрофных микроорганизмов Института экологии и генетики микроорганизмов (официальный акроним коллекции ИЭГМ, номер во Всемирной федерации коллекций культур 768) [7], принадлежащие к видам Rhodococcus erythropolis (21 штамм) и R. ruber (8 штаммов). В отдельных экспериментах использовали мутантные клоны R. ruber ИЭГМ 231, полученные методом неспецифического in vivo Tn5-мутагенеза [3]. Для получения УФ мутантов клетки R. ruber ИЭГМ 381 выращивали в питательной среде (г/л): (NH4)2SO4 – 2,0; K2HPO4 – 2,0; MgSO4 – 0,1; CaCl2×2H2O – 0,01; FeSO4×7H2O – 0,01; глицерин – 10,0; дрожжевой экстракт – 4,0. Через 48 ч клетки стерильно отделяли от среды центрифугированием (15 мин, 3000 об/мин), трижды промывали фосфатно-щелочным буфером (KH2PO4/NaOH) pH 7,0. Полученную суспензию (OП600 2,5) облучали с помощью УФ лампы ДРТ-240 (Россия) в течение 5 мин. Химический мутагенез клеток R. ruber ИЭГМ 233, предварительно выращенных в мясопептонном бульоне и промытых ацетатным буфером pH 4,5, проводили в течение 10 мин при 28 ºС в 0,05 М растворе азотистой кислоты (HNO2), затем для остановки реакции добавляли среду 1ХА [5]. Колонии УФ и HNO2 мутантов получали после высева клеточных суспензий на чашки Петри с мясопептонным агаром и инкубации в течение 24 ч при 28 °С.
Родококки выращивали в условиях периодического культивирования на орбитальной качалке Certomat IS «Sartorius» (Германия) (150 об/мин) при 28 °С. Базовый состав минеральной среды включал следующие компоненты (г/л): KNO3 – 1,0; KH2PO4 – 1,0; K2HPO4×3H2O – 1,0; NaCl – 1,0; MgSO4×7H2O – 0,2; СaCl2×2H2O – 0,02. В среду добавляли 1 г/л дрожжевого экстракта, 0,1 об. % раствора микроэлементов по Постгейту и 0,7 г/л н-гексадекана. В исследованиях по подбору оптимальных условий биотрансформации β-ситостерола в высоких концентрациях в среду вносили 1,2 г/л н-гексадекана и 0,015 г/л пальмитиновой кислоты. β-Ситостерол добавляли в инкубационную среду одновременно с инокулятом или через 48 ч роста родококков в концентрации 0,5; 2,0; 6,0 или 10,0 г/л в виде 10 % раствора в изопропаноле или смеси с β-циклодекстрином (1:1) в 1,2 % водном растворе Твина-80. Исследования по биотрансформации β-ситостерола нерастущими клетками проводили в фосфатно-щелочном буфере (KH2PO4/NaOH) pH 6, 7 или 8.
В качестве посевного материала использовали родококки (5,0×105 клеток/мл), выращенные на мясопептонном агаре и отобранные в экспоненциальной фазе роста. Эксперименты по биотрансформации β-ситостерола нерастущими клетками родококков проводили с использованием бактериальных культур, выращенных на агаризованной среде состава (г/л): K2HPO4 – 1,0; (NH4)2HPO4 – 1,5; MgSO4 × 7H2О – 0,1; FeSO4 × 7H2О – 0,01; ZnSO4 × 7H2О – 0,002; глюкоза – 5,0, дрожжевой экстракт – 10,0 с добавлением 0,2 г/л β-ситостерола в виде 10 % раствора в изопропаноле.
В работе применяли реактивы российских производителей марки «х.ч.» или «ч.д.а.» и фирмы «Sigma-Aldrich» (США). В экспериментах использовали β-ситостерол 70,0 % чистоты «Sigma-Aldrich» (США). Образец стигмаст-4-ен-3-она получен из Новосибирского института органической химии им. Н.Н. Ворожцова СО РАН, ацетат β-ситостерола синтезировали как описано ранее [1]. Продукты микробного окисления β-ситостерола экстрагировали этилацетатом (3×50 мл). Объединенные экстракты высушивали над Na2SO4, растворитель удаляли на роторном испарителе. Качественный и количественный состав полученных смесей анализировали методом тонкослойной хроматографии с использованием пластин «Sorbfil» марки ПТСХ-АФ-А-УФ фирмы ЗАО «Сорбполимер» (Россия) [4], УФ-спектроскопии на спектрофотометре Lambda EZ201 «Perkin-Elmer» (США) и хроматомасс-спектрометрической системы Agilent 6890/5973N (кварцевая колонка HP-5MS SN US 15189741-1) «Agilent technology» (США). Эксперименты проводили в 3-кратной повторности. Статистическую обработку результатов осуществляли с использованием программы Excel 2003, рассчитывая среднее арифметическое и стандартную ошибку.
Результаты исследования и их обсуждение
По нашим данным, представители R. ruber эффективно (55–98 %) трансформируют β-ситостерол (0,5 г/л) с образованием стигмаст-4-ен-3-она при условии добавления исходного стерола через 2 суток роста бактериальных клеток в среде с н-гексадеканом (рис. 1, А). Генетическая модификация штаммов, обладающих в присутствии н-гексадекана низкой стеролтрансформирующей активностью, методом неспецифического in vivo Tn5-мутагенеза позволяет значительно повысить их способность к биотрансформации β-ситостерола. Получены мутантные клоны Tn5-11, Tn5-25, Tn5-29, Tn5-37, катализирующие 60 % конверсию β-ситостерола в стигмаст-4-ен-3-он, что в 4 раза выше таковой исходной культуры R. ruber ИЭГМ 231 (рис. 1, Б).
В отдельных экспериментах установлено, что внесение в среду с н-гексадеканом 0,15 г/л пальмитиновой кислоты в качестве индуктора ферментативной активности представителей вида R. erythropolis обеспечивает эффективное получение стигмаст-4-ен-3-она при добавлении β-ситостерола одновременно с инокулятом. В результате исследования способности родококков трансформировать β-ситостерол в концентрации 2,0 г/л выявлено, что степень образования стигмаст-4-ен-3-она при использовании Твина-80 в качестве растворителя исходного стерола в среднем составляет 19 %. Лишь отдельные штаммы R. erythropolis (ИЭГМ 10 и ИЭГМ 487, в частности) катализируют 44,9–49,5 % конверсию β-ситостерола в целевой продукт. Более высокая степень биотрансформации β-ситостерола достигается в условиях его добавления в виде смеси с β-циклодекстрином (β-ЦД) и Твином-80, при этом каталитическая активность штаммов R. erythropolis, как правило, в 1,1–6,0 раз превышает таковую R. ruber. Максимальная (77,3–79,1) степень биотрансформации β-ситостерола в стигмаст-4-ен-3-он регистрируется при использовании R. erythropolis ИЭГМ 179 и ИЭГМ 487 (рис. 2).
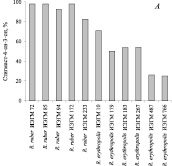
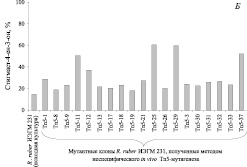
Рис. 1. Биотрансформация β-ситостерола в стигмаст-4-ен-3-он в присутствии н-гексадекана в условиях внесения β-ситостерола через 2 суток роста родококков. А – нативными штаммами, Б – мутантными клонами R. ruber ИЭГМ 231
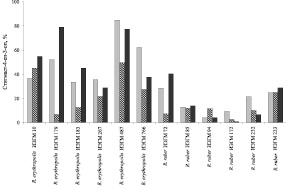
Рис. 2. Биотрансформация β-ситостерола родококками в присутствии н-гексадекана (0,25 %) и пальмитиновой кислоты (0,015 %) в условиях добавления 0,5 г/л исходного стерола в 5 мл изопропанола ( ); 2,0 г/л в 20 мл Твина-80 (
); 2,0 г/л в 20 мл Твина-80 ( ); 2,0 г/л в виде смеси с β-ЦД (1:1) в 40 мл 1,2 % водного раствора Твина-80 (
); 2,0 г/л в виде смеси с β-ЦД (1:1) в 40 мл 1,2 % водного раствора Твина-80 ( ). β-Ситостерол вносили в инкубационную среду одновременно с инокулятом. Приведены данные после 5 сут культивирования родококков
). β-Ситостерол вносили в инкубационную среду одновременно с инокулятом. Приведены данные после 5 сут культивирования родококков
Установлено, что методы химического и УФ-мутагенеза не способствуют повышению стеролтрансформирующей активности представителей R. ruber. Так, степень образования стигмаст-4-ен-3-она мутантными клонами R. ruber ИЭГМ 233 и ИЭГМ 381 в условиях добавления 2,0 г/л β-ситостерола снижается в 2,4–5,3 раза по сравнению с исходными культурами (таблица).
Дальнейшие эксперименты проводили с использованием представителей R. erythropolis, наиболее эффективно трансформирующих β-ситостерол в концентра ции 2,0 г/л.
Биотрансформация β-ситостерола мутантными клонами R. ruber
|
Химический мутагенез |
УФ мутагенез |
||
|
Мутантый клон |
Стигмаст-4-ен-3-он, % |
Мутантый клон |
Стигмаст-4-ен-3-он, % |
|
R. ruber ИЭГМ 233 (исходная культура) |
28,7 ± 8,34 |
R. ruber ИЭГМ 381 (исходная культура) |
20,3 ± 7,92 |
|
233–9 |
10,4 ± 4,75 |
381–1 |
8,5 ± 3,14 |
|
233–11 |
5,8 ± 2,16 |
381–9 |
8,2 ± 2,08 |
|
233–14 |
5,1 ± 1,44 |
381–10 |
8,1 ± 1,97 |
|
233–15 |
8,6 ± 3,81 |
381–16 |
7,5 ± 0,84 |
|
233–19 |
5,4 ± 1,32 |
381–18 |
8,2 ± 2,43 |
Примечание: Биотрансформацию β-ситостерола проводили в присутствии н-гексадекана (0,25 %) и пальмитиновой кислоты (0,015 %) в условиях добавления 2 г/л исходного стерола в виде смеси с β-ЦД (1:1) в 40 мл 1,2 % водного раствора Твина-80.
Установлено, что в условиях добавления 6,0 г/л исходного стерола в виде смеси с β-ЦД и Твином-80 штаммы R. erythropolis проявляют, как правило, высокую (более 90 %) стеролтрансформирующую способность (рис. 3, А). При повышении концентрации β-ситостерола до 10 г/л активность большинства исследуемых культур не превышает 60 % (рис. 3, А, Б).
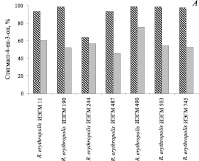
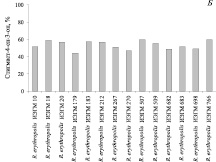
Рис. 3. Биотрансформация β-ситостерола представителями R. erythropolis в условиях добавления 6( ) или 10 (
) или 10 ( ) исходного стерола в виде смеси с β-ЦД (1:1) в 1,2 % водном растворе Твина-80. Приведены данные после 5 суток культивирования родококков
) исходного стерола в виде смеси с β-ЦД (1:1) в 1,2 % водном растворе Твина-80. Приведены данные после 5 суток культивирования родококков
Наиболее высокий (75,4–98,9 %) уровень конверсии β-ситостерола в стигмаст-4-ен-3-он достигается при использовании R. erythropolis ИЭГМ 490.
Физиологическое состояние бактериальных клеток может оказывать значительное влияние на процесс биотрансформации органических соединений, в связи с этим нами проведены эксперименты по оценке способности нерастущих клеток родококков к биотрансформации b-ситостерола. С использованием культур R. erythropolis ИЭГМ 487 и R. ruber ИЭГМ 233, обладающих в присутствии н-гексадекана высокой стеролтрансформирующей активностью, установлено, что в процессе инкубации бактерий в фосфатно-щелочном буфере с добавлением b-ситостерола (2,0 г/л) в качестве продукта трансформации исходного стерола обнаруживается ацетат b-ситостерола. Максимальная (75 %) степень образования данного продукта регистрируется в кислой реакции среды при использовании клеток R. ruber ИЭГМ 233, предварительно выращенных в глюкозосодержащей среде с добавлением β-ситостерола в качестве индуктора.
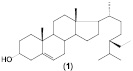
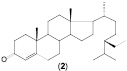
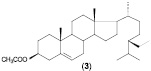
Таким образом, в результате проведенных исследований определены основные особенности процесса биотрансформации β-ситостерола (1) нативными и мутантными штаммами родококков с образованием стигмаст-4-ен-3-она (2). Детально исследована способность R. erythropolis к биотрансформации исходного стерола в высоких (2, 6 или 10 г/л) концентрациях. Проведена оценка способности нерастущих клеток родококков к биотрансформации β-ситостерола, при этом в качестве продукта реакции обнаружен ацетат β-ситостерола (3).
Работа выполнена при финансовой поддержке грантов РФФИ и Министерства образования и науки Пермского края (проект № 14-04-69005-р_урал_а).
Рецензенты:
Вихарева Е.В., д.фарм.н., профессор, заведующая кафедрой аналитической химии, ФГБОУ ВПО «Пермская государственная фармацевтическая академия», г. Пермь;
Володин В.В., д.б.н., профессор, заведующий лабораторией биохимии и биотехнологии растений, Институт биологии Коми НЦ УрО РАН, г. Сыктывкар.
Работа поступила в редакцию 05.09.2014.
Библиографическая ссылка
Ноговицина Е.М., Гришко В.В., Бажутин Г.А. ТРАНСФОРМИРУЮЩАЯ АКТИВНОСТЬ НАТИВНЫХ И МУТАНТНЫХ ШТАММОВ РОДОКОККОВ В ОТНОШЕНИИ β-СИТОСТЕРОЛА // Фундаментальные исследования. 2014. № 9-10. С. 2218-2222;URL: https://fundamental-research.ru/en/article/view?id=35300 (дата обращения: 16.07.2026).