


Scientific journal
Fundamental research
ISSN 1812-7339
"Перечень" ВАК
ИФ РИНЦ = 1,798
BIOECOLOGICAL ASPECTS OF SUCCESSION INVESTIGATION OF A HAY INFUSION MICROCOSM

Сукцессия (от лат. successio - преемственность, наследование) - это процесс саморазвития сообщества. Экологическую сукцессию можно определить по следующим параметрам: развитие видовой структуры во времени, изменение условий среды обитания под влиянием предшествующих организмов, смена видов в экосистеме, которая контролируется сообществом, и происходящие в системе обменные процессы. Кульминацией развития является формирование стабильного сообщества, длительность существования которого определяется поступлением (или отсутствием) энергии из внешней среды [2, 5].
Сукцессия идет в направлении фундаментального сдвига потока энергии в сторону увеличения ее количества, направленного на поддержание системы в соответствии с правилом максимума энергии поддержания зрелой системы, сформулированным Г. Одумом и Р. Пинкертоном [7].
Актуальность проведенной работы заключается в том, что получаемые в культуре сенного настоя простейшие гетеротрофы могут быть использованы как тест-организмы для оценки токсичности водной среды.
Однако, как показывает практика, получение культур сенного настоя требует проведения предварительных исследований, так как скорость сукцессионных процессов, особенно на ранних стадиях, высока, смена видов может произойти очень быстро, и не всегда понятны причины, вызывающие гибель или исчезновение организмов. Поэтому для длительного поддержания их жизнедеятельности необходимо изучить оптимальные условия существования организмов, выбранных в качестве тест-объектов.
Различают автотрофные и гетеротрофные сукцессии. В отличие от автотрофной сукцессии длительность гетеротрофной сукцессии определяется количеством энергии, находящейся в мертвой органике сенного настоя. Сенной настой, называемый в экологии микрокосмом, служит хорошей моделью сукцессии, которая в природе происходит, например, в гниющих стволах деревьев, в трупах животных, в фекалиях и на вторичных стадиях очистки сточных вод.
Цель исследования
Проанализировать особенности сериальных стадий гетеротрофной сукцессии в культуре сенного настоя.
Задачи исследования
1) изучить видовой состав сообщества в ходе эксперимента;
2) построить сукцессионный ряд, отражающий закономерности развития модельного микрокосма.
Материал и методы исследования
Основу материалов составили результаты серий наблюдений за динамикой развития культуры сенного настоя. Приготовление сенного настоя осуществлялось согласно апробированным методикам [1, 4, 8]. Периодически сенной настой подкармливали несколькими каплями стерилизованного молока. По мере испарения в сенной настой доливали отстоянную водопроводную воду. Наблюдения за микроорганизмами проводили в течение 4-х месяцев. Подсчет осуществлялся под микроскопом в 10 полях зрения при увеличении: об.2х, ок.8х; об.20х, ок.10х. Идентификацию простейших проводили при увеличении: об.40х, ок.10х. Пробы для просмотра отбирали с поверхности, в толще воды и у дна. Статистическая обработка результатов проводилась стандартными методами.
Работа выполнена на кафедре биоэкологии МГТУ в рамках ГБ НИР «Разработка методов биоиндикации и биотестирования пресноводных водоемов Кольского полуострова».
Результаты исследования и их обсуждение
Как показали наблюдения, основными факторами, лимитирующими рост микроорганизмов в микрокосме сенного настоя, являются питательный субстрат, газовый режим, количество и возраст цист в исходном материале.
Через двое суток в сосуде с настоем появляется бактериальная пленка. Просмотр проб показал наличие в среде многочисленных бактериальных клеток, жгутиконосцев, множество цист разного размера. Из простейших организмов первыми появились ресничные инфузории Colpoda steini и Aspidisca turrida. Основная масса инфузорий С. steini находилась у дна. Численность инфузорий по трем горизонтам составила 4,1; 2,0; 7,2 экз./п.зр. микроскопа соответственно.
Спустя 10 суток число С. steini стало уменьшаться и составило 1,7; 2,1; 1,3 экз./п.зр. микроскопа. Решающим фактором для инфузорий (в целом для всех простейших) является пища. Известно, что С. steini живут в среде, бедной растворенной органикой и при возрастании органики они инцистируются (рис. 1). Colpoda в течение 12 часов успевают эксцистироваться, обеспечить себя пищей, разделиться и снова инцистироваться [3].
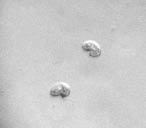

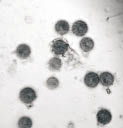
Рис. 1. Сенной настой: этапы инцистирования инфузории Colpoda steini
Одновременно в пробах были обнаружены единичные экземпляры двух видов брюхоресничных инфузорий, принадлежащих к роду Stylonychia - это подвижные, вооруженные щетинками и пучками ресничек (цирри) организмы (рис. 2а). Периодически в сенной настой в качестве пищи добавляли несколько капель стерилизованного молока. Добавление 20‒25 капель стимулировало размножение стилонихий. Подкормка в количестве 30 капель привела к инцистированию инфузорий. С увеличением количества стилонихий исчезали равноресничные инфузории.
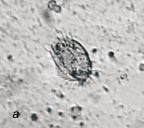
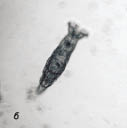
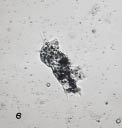
Рис. 2. Сенной настой:
а - инфузория Stylonychia sp.; б - коловратка Philodina sp.;
в - амеба фагоцитирует цисты инфузорий
Спустя 1,5 месяца число стилонихий возросло и составило 3; 2; 5 экз./п.зр. микроскопа. Вслед за стилонихиями в пробах появились коловратки (рис. 2б). Отмечено несколько видов коловраток рода Philodina, Rotaria и Dissotrocha. При наступлении неблагоприятных условий коловратки способны впадать в состояние анабиоза по типу ангидробиоза или высушивания. Численность голых амеб в пробе постепенно увеличивалась и составила 5 экз./п.зр. микроскопа. Они активно фагоцитируют, часто внутри амеб можно увидеть цисты инфузорий (рис. 2в).
В эти сроки в пробах были обнаружены ресничные черви семейства Stenostomidae, первоначально в небольшом количестве, их численность с каждым днем увеличивалась в геометрической прогрессии.
Через 2 месяца численность ресничных червей достигла 200 экз. в объеме 4 мл пробы. В этот период отмечено появление единичных нематод, численность которых быстро возрастала. Таким образом, в настое доминировали ресничные черви и нематоды (рис. 3).
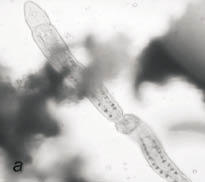
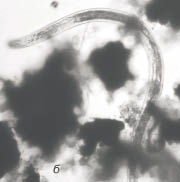
Рис. 3. Сенной настой:
а - ресничный червь, б - нематода
Изучаемый микрокосм можно отнести к частично закрытой системе, в которой происходит газообмен с атмосферой. К внешнему воздействию на систему можно отнести периодическое добавление стерилизованного молока в качестве пищи и отстоянной водопроводной воды.
По мере минерализации мертвой органики происходит осветление воды и снижение источников энергии для гетеротрофов.
Через 3 месяца число видов в микрокосме снижается. Численность нематод, ресничных червей и амеб составила 66; 20; 1 экз./п.зр. микроскопа соответственно.
Спустя 4 месяца в исследуемом микрокосме исчезли все инфузории, коловратки, значительно снизилось количество ресничных червей и нематод. Однако преобладали нематоды. Средняя численность ресничных червей и нематод составила 10 и 45 экз. в объеме 4 мл пробы.
Таким образом, поставленный эксперимент по получению культуры простейших гетеротрофов позволил установить сукцессионный ряд организмов, появляющихся в определенной последовательности: бактерии → жгутиконосцы → голые амебы → равноресничные инфузории → брюхоресничные инфузории → коловратки → ресничные черви → нематоды. Численность вышеперечисленных представителей микроорганизмов варьируется, они в том или ином количестве присутствуют почти постоянно [6]. Однако можно выделить периоды доминирования инфузорий, коловраток, амеб, червей, нематод.
Описанный со стилонихиями факт показал, что эти инфузории могут инцистироваться при изобилии пищи. Ресничные черви и нематоды активны и быстро размножаются при обилии органики. Их численность снижается по мере деструкции органического вещества в микрокосме.
Заключение
Как показали серии наблюдений, полученный сукцессионный ряд не является абсолютным. Важными факторами, определяющими состав сообщества, являются органические вещества, поступающие с сенным настоем и пищей, количество цист в почве, газовый режим среды, продукты метаболизма и др.
По мере развития сукцессии видовое разнообразие увеличивается, меняется видовой состав сообщества, усложняются и удлиняются жизненные циклы, увеличивается размер организмов, развиваются взаимовыгодные симбиозы, усложняется трофическая структура экосистемы. Однако пик разнообразия приходится вовсе не на стадию климакса, а либо на середину сукцессионного ряда, либо ближе к климаксу.
Таким образом, в процессе сукцессии происходит, по-видимому, переход от количества продукции к качеству, сопровождаемый генетическими изменениями, охватывающими все сообщество.
По мере усложнения трофической структуры микрокосма увеличивается время оборота биогенных веществ. Кроме того, циклы многих биогенных элементов, таких как азот, фосфор и кальций, замыкаются в пределах данной экосистемы. Все это способствует накоплению биогенных элементов, то есть, в процессе сукцессии количество этих элементов на входе оказывается большим, чем их количество на выходе. В зрелой климаксной системе вход и выход биогенных элементов почти уравновешивается, однако некоторое накопление все же происходит. Таким образом, равновесная система более независима от внешних поступлений вещества.
После достижения климакса динамические процессы не прекращаются, но развитие микрокосма замедляется. Одни организмы сменяются другими, происходит внедрение в экосистему новых видов, формируются новые экологические ниши и симбиотические связи, совершенствуются адаптации организмов к экологическим факторам.
Формирование экосистемы сенного настоя полностью соответствует установленным теоретическим предпосылкам. Экосистему формирует биотический фактор - виды ранних сукцессионных стадий меняют условия существования и подготавливают почву для последующих поселенцев; первые виды противостоят вторжению других видов (ингибируют их) и сохраняются до тех пор, пока не будут вытесненными в результате конкуренции, выедания гетеротрофами, самоотравления продуктами своей жизнедеятельности и т.п.
Среди организмов, полученных в лабораторном микрокосме, стабильно поддерживают свою численность инфузория рода Stylonychia и ресничный червь семейства Stenostomidae, что является одним из главных условий использования их в качестве тест-объектов.
Список литературы
- Биологический контроль окружающей среды. Биоиндикация и биотестирование: учеб. пособие для вузов / О.П. Мелехова [и др.]; под ред. О.П. Мелеховой и Е.И. Егоровой. - М. : Академия, 2007. - 288 с.
- Биотестирование и прогноз изменчивости водных экосистем при антропогенном загрязнении / Г.Г. Матишов [и др.]. - Апатиты: Изд-во КНЦ РАН, 2003. - 468 с.
- Дажо Р. Основы экологии / пер. с фр. В.И. Назарова; ред. В.В. Алпатова. - М.: Прогресс, 1975. - 415 с.
- Микулин А.Е. Живые корма // Клуб натуралистов «Дельфин». - М.: Дельфин, 1994. - 104 с.
- Одум Юджин П. Экология: в 2 т. / пер. с англ. Ю.М. Фролова; под ред. В.Е. Соколова. - М.: Мир, 1986. - Т. 1. - 328 с.
- Пахомова Н.А., Минченок Е.Е. Модель гетеротрофной сукцессии в культуре сенного настоя // Наука и образование - 2010 [Электронный ресурс]: юбилейная междунар. науч.-техн. конф., посвящ. 60-летию МГТУ, Мурманск, 5‒9 апр. 2010 г. / Федер. агентство по рыболовству, Мурман. гос. техн. ун-т [и др.]. - Мурманск, 2010. - С. 834-836. - URL: http://www.mstu.edu.ru/ (дата обращения: 10.12. 2010).
- Реймерс Н. Ф. Экология: Теории, законы, правила, принципы и гипотезы. - М.: Россия Молодая, 1994. - 366 с.
- Федорова А.И., Никольская А.Н. Практикум по экологии и охране окружающей среды: учеб. пособие для вузов. - М.: ВЛАДОС, 2001. - 288 с.
Рецензенты:
Перетрухина А.Т., д.б.н., профессор, зав. кафедрой микробиологии Мурманского государственного технического университета, Мурманск;
Макаревич П.Р., д.б.н., профессор, первый зам. директора по науке Мурманского морского биологического института, Мурманск.
Библиографическая ссылка
Пахомова Н.А, Минченок Е.Е БИОЭКОЛОГИЧЕСКИЕ АСПЕКТЫ ИЗУЧЕНИЯ СУКЦЕССИИ В МИКРОКОСМЕ СЕННОГО НАСТОЯ // Фундаментальные исследования. 2011. № 5. С. 201-205;URL: https://fundamental-research.ru/en/article/view?id=21284 (дата обращения: 16.07.2026).