



Многочисленные научные обзоры отражают тот факт, что оксид азота (NO) играет ключевую роль в регуляции кардиоваскулярного гомеостаза: NO вовлекается в механизмы гемостаза, контролирует барорефлекс, оказывает стресс-лимитирующие эффекты, регулирует баланс вегетативных влияний на сердечно-сосудистую систему [5,9]. В наших предыдущих исследованиях было показано, что снижение генерации NO ассоциируется с повышением кардиоваскулярной стресс-реактивности и развитием артериальной гипертонии [1,3,6].
Таким образом, количественное описание сложности эволюционной динамики сигналов кровяного давления и сердца в условиях изменения активности NO-ергической системы при различных физиологических состояниях организма могут иметь важное прогностическое и диагностическое значение.
Цель работы
Изучение эффектов активности NO-синтазы на динамику кровяного давления у самок крыс на основе применения теории вейвлетов. Этот выбор обусловлен результатами предыдущих исследований [6], которые позволили сделать предварительные выводы о том, что критические изменения в деятельности ССС, наблюдаемые у самок в условиях блокады синтеза оксида азота, сочетаются у них с уменьшением сложности динамики в высокочастотной области спектра (ВЧ, 0,75-3,0 Гц) на фоне менее выраженных изменений динамики в низкочастотной области (НЧ, 0,25-0,75 Гц). Напротив, у самцов, сердечно-сосудистая система которых менее чувствительна к блокаде NO-ергического звена регуляции, не наблюдалось значительных изменений сложности ни в ВЧ-, ни НЧ-областях спектра.
Материалы и методы исследования
Эксперименты были выполнены на 24 белых половозрелых самках крыс. Животным были имплантированы внутрисосудистые катетеры под нембуталовым наркозом (45 мг/кг) для прямого измерения кровяного давления (КД). Запись сигналов кровяного давления осуществляли с помощью совместимого с компьютером многоканального измерительно-вычислительного комплекса PowerLab/400 ML401 и программного обеспечения Chart 4 (ADInstruments Ltd., Австралия). Блокаду активности NO-синтазы проводили путем введения NG-nitro-L-arginine-methyl-ester (L-NAME, 10 мг/кг, Sigma Chemical Co, iv). Все хирургические манипуляции проводили под общей нембуталовой анестезией (0,35 мг/кг, ip). Для статистической обработки экспериментальных данных использовали пакет программ Statistica 5.0. Исследование структуры записанных сигналов проводилось на основе вейвлет-анализа [2,4], который является в настоящее время одним из наиболее популярных подходов к изучению эволюционной динамики физиологических процессов. К числу его достоинств относится возможность изучать как стационарные, так и нестационарные (переходные) процессы в динамике систем различной природы, возможность идентифицировать мгновенные характеристики многочастотных колебательных процессов [7] и т.д. Данный метод традиционно называют «математическим микроскопом» [2], который позволяет детально изучить структуру сложных сигналов в широком диапазоне масштабов.
Математическая процедура непрерывного вейвлет-преобразования некоторого сигнала предполагает его разложение по системе базисных функций, которые формируются путем масштабных преобразований и переносов вдоль временной оси так называемого «материнского» вейвлета ψ:
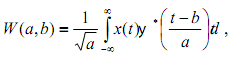
где x(t) - анализируемый сигнал КД, W(a,b) - коэффициенты преобразования, a - масштаб наблюдения, b - параметр смещения вдоль оси времени. При проведении частотно-временного анализа экспериментальных данных традиционно используют вейвлет Морле:
![]()
Параметр f0 данной функции подбирается таким образом, чтобы достичь компромисса между разрешением сигнала x(t) по времени и по частоте [2]. Вместо коэффициентов W(a,b) на практике удобно оперировать с плотностью энергии E(a,b)=|W(a,b)|2, эволюция локальных максимумов которой соответствует мгновенным частотам ритмических процессов, а усреднение плотности энергии по времени является аналогом сглаженного спектра мощности. Примеры вейвлет-анализа сигналов КД можно найти в работе [6]. В результате проведения данного анализа идентифицируются временные зависимости мгновенных частот f1(t) и f2(t), характеризующих динамику ритмических процессов в НЧ- и ВЧ-областях спектра соответственно.
Для количественного описания сложности динамики КД будем использовать одну из стандартных характеристик - энтропию Шеннона:
![]()
где N - число разбиений диапазона изменения мгновенной частоты, Pk - вероятность того, что значение мгновенной частоты попадает в k-й интервал разбиения ширины Δ. По аналогии с работой [6] для анализируемого ритма f1(t) и f2(t) всех экспериментальных данных целесообразно использовать одинаковую меру неопределенности Δ.
Таким образом, в рамках проводимого исследования мы акцентируем внимание на отслеживании временной эволюции различных ритмических составляющих (мгновенных частот, соответствующих тому или иному ритму) и количественном описании сложности эволюционной динамики применительно к каждому ритму в отдельности. В общем случае для постоянного значения частоты (стабильный ритм) Hp=0 при совершенно случайном поведении мгновенной частоты ритма Hp принимает максимальное значение, равное log N.
Результаты исследования показали, что блокада синтеза NO сопровождалась уменьшением значений энтропии Hp, что свидетельствует об эффекте стабилизации частоты ритмических процессов, которые диагностируются как в низкочастотной, так и в высокочастотной областях спектра. В частности, они четко диагностировались в высокочастотной области, изменения количественной меры сложности в которой в отдельных экспериментах достигает 10-15% при блокаде синтеза NO по сравнению с базовым уровнем. В низкочастотной области также можно обнаружить изменения сложности экспериментальных данных, связанные с введением L-NAME, но они обычно менее выраженные (не более 10%). Однако как в НЧ-, так и в ВЧ-областях эффекты стабилизации ритмической динамики являются значительными в соответствии с критерием Манна-Уитни (p<0,05). Данный эффект согласуется с выводами работы [8], проиллюстрировавшей влияние L-NAME на различные механизмы почечной регуляции кровотока.
Результаты статистического анализа по всем экспериментальным данным свидетельствуют о том, что самки демонстрируют существенные изменения сложности динамики, обусловленные блокадой синтеза NO в высокочастотной области (4,85±0,06 для базового уровня, 4,68±0,15 при блокаде). В области низкочастотной динамики было зафиксировано менее существенное снижение меры сложности Hp - со значения 3,42±0,05 (базовый уровень) до 3,34±0,12 (блокада синтеза NO). На рисунке для наглядности представлены результаты расчета нормированных значений Hp, когда за единичный уровень принимается среднее значение энтропии Шеннона, соответствующее базовому уровню.
Наряду с применением описанного подхода нами также проводилась количественная оценка изменений в динамике ССС, вызванных блокадой синтеза NO, с помощью стандартных физиологических параметров, таких, как среднее артериальное давление (ср.АД) и частота сердечных сокращений (ЧСС). Согласно результатам предыдущей работы [6], данные характеристики свидетельствуют о том, что самки крыс более чувствительны к блокаде синтеза NO по сравнению с самцами. Было обнаружено, что L-NAME вызывает увеличение ср.АД у самок на 25% и уменьшение ЧСС на 18%.
Предлагаемый подход на основе совместного применения теории вейвлетов и расчета количественных критериев сложности динамики позволил нам исследовать ряд важных особенностей процессов адаптации, таких как длительность переходных процессов в динамике ССС. Отметим, что эта длительность может различаться для динамики в разных диапазонах спектра вариабельности сердечного ритма и для ЧСС.
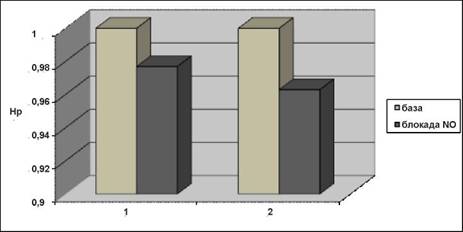
Примечание: Изменение сложности динамики КД самок крыс при блокаде синтеза NO, Цифрами 1 и 2 обозначена динамика в НЧ- и ВЧ-областях спектра соответственно. Результаты представлены в нормированных единицах.
Заключение
Проведенные исследования позволяют сделать вывод о том, что блокада синтеза NO сопровождается стабилизацией динамики кровяного давления, ассоциирующейся как с низкочастотной, так и (в большей степени) с высокочастотной областью. Отметим, что, с одной стороны, существуют доказательства влияния эндотелиального уровня NO на динамику в низкочастотной области, но, с другой стороны, известно, что различные физиологические эффекты фармакологической блокады NO зависят от типа блокатора и его концентрации. В данной работе применялся L-NAME, который является неспецифическим блокатором NO. Кроме того, использовалась доза 10 мг/кг, которая полностью блокирует синтез NO. Полученные результаты согласуются с результатами анализа почечного кровотока [8], продемонстрировавшими эффекты L-NAME на разные механизмы регуляции кровяного давления. Обнаруженный эффект стабилизации динамики сигналов кровяного давления самок крыс ассоциируется у них с высокой кардиоваскулярной устойчивостью к стрессу и развитию артериальной гипертонии.
Список литературы
- Анищенко Т.Г., Глушковская-Семячкина О.В., Бердникова В.А., Синдякова В.А. Половые особенности кардиоваскулярной стресс-реактивности у здоровых и гипертензивных крыс // Бюллетень экспериментальной биологии и медицины. - 2007. - Т.143, № 2. - С.136-140.
- Астафьева Н.М. Вейвлет-анализ: основы теории и примеры применения // Успехи физических наук. - 1996. - T. 166, № 11. - С. 1145-1168.
- Глушковская-Семячкина О.В., Анищенко Т.Г., Синдякова Т.А. Половые различия в содержании оксида азота у здоровых и гипертензивных крыс в условиях покоя и стресса // Бюллетень экспериментальной биологии и медицины. - 2006. - Т.142, № 8. С. 13-17.
- Grossmann A., Morlet J. Decomposition of hardy functions into square integrable wavelets of constant shape // S.I.A.M. J. Math. Anal. - 1984. - Vol. 15. - P. 723-736.
- Orshal J., Khalil R. Gender, sex hormones, and vascular tone // Am J Physiol Regul Integr Comp Physiol. - 2004. - Vol. 286. - P. R233-R249.
- Pavlov A.N., Anisimov A.A., Semyachkina-Glushkovskaya O.V., Matasova E.G., Kurths J. Analysis of blood pressure dynamics in male and female rats using the continuous wavelet-transform // Physiological Measurement. - 2009. - Vol. 30. - P. 707-717.
- Pavlov A.N., Sosnovtseva O.V., Pavlova O.N., Mosekilde E., Holstein-Rathlou N.-H. Characterizing multimode interaction in renal autoregulation // Physiological Measurement. - 2008. - Vol. 29. - P. 945-958.
- Sosnovtseva O.V., Pavlov A.N., Pavlova O.N., Mosekilde E., Holstein-Rathlou N.-H. Characterizing the effect of L-name on intra- and inter nephron synchronization // European Journal of Pharmaceutical Sciences. - 2009. - Vol. 36. - P. 39-50.
- Toda N., Okamura T. The pharmacology of nitric oxide in the peripheral nervous system of blood vessels // Pharmacol Rev. - 2003. - Vol. 55. - Р. 271-324.