



Значительное влияние на микробиоценоз кишечника рыб оказывают вид и биохимические характеристики кормовых объектов [10], в связи с этим качественный и количественный состав составляющих его микроорганизмов различен у рыб с различным типом питания [4]. Всеядные виды рыб обладают большим микробным разнообразием, чем хищные [12].
Байкальский омуль Coregonus migratorius (Georgi, 1775) и черный байкальский хариус Thymallus baicalensis отличаются по пищевым стратегиям и принадлежат к разным (пелагической и бентической) трофическим сетям озера Байкал. Известны три морфоэкологические группы (МЭГ) байкальского омуля (пелагическая, прибрежная и придонно-глубоководная), занимающие различные пищевые ниши. В зимне-весенний период омуль всех МЭГ имеет сходные пищевые спектры (Epischura baicalensis и Macrohectopus branickii, молодь рыб, донные амфиподы), т.к. обитает в придонных слоях склоновой зоны озера. В мае-июне омуль расходится по различным биотопам: придонно-глубоководный остается в придонных слоях воды на значительных глубинах (сезонные изменения рациона не выражены); пелагический нагуливается в пелагиали и в придонных слоях воды (сезонные изменения незначительны); омуль прибрежной МЭГ обитает в прибрежной зоне озера (кормовая база крайне изменчива сезонно и территориально) [11].
Черный байкальский хариус преимущественно ведет образ жизни, связанный с речными условиями. В течение всего года он образует плотные скопления в литорали Байкала в предустьевых участках притоков, в которых происходит его нерест и частично нагул [3]. По характеру питания черного байкальского хариуса можно считать эврифагом, т.к. спектр потребляемых им организмов очень широк и включает в себя как животных бентоса (личинок ручейников, амфипод, гастропод и др.), так и имаго амфибиотических насекомых [6, 7].
Цель настоящего исследования – провести сравнительный анализ генетического разнообразия кишечных микробиоценозов черного байкальского хариуса и байкальского омуля, лососевидных рыб с разной пищевой стратегией и распределением в озере.
Материал и методы исследования
Материалом для работы послужили два вида лососевидных рыб озера Байкал: черный байкальский хариус (27 экз.) и байкальский омуль (67 экз.). Сборы материалов по акватории озера Байкал проводили в период открытой воды (июнь и сентябрь) разноглубинным тралом РК-15/30. Для сбора черного байкальского хариуса использовали жаберные сети (ячея 14–24 мм), которые выставлялись на глубинах 0,5–8,0 м в мае 2008 г. в районе б. Песчаная (южная котловина озера Байкал). Непосредственно после отлова в лабораторных условиях взяты фрагменты кишечного тракта (КТ) рыб.
Молекулярно-генетический анализ проводили согласно адаптированным ранее методикам [2]. Структуру микробных сообществ кишечной микробиоты байкальского омуля трех МЭГ и черного байкальского хариуса исследовали с помощью групп-специфичной ПЦР [1]. Ампликоны, полученные индивидуально для каждой особи на консервативных бактериальных праймерах [2], объединяли для анализируемых групп и суммарные образцы, лигировали с использованием набора GeneJETTM PCR Cloning Kit (Fermentas, Литва). Все выросшие колонии анализировали методом ПЦР с праймерами, рекомендованными производителем (Fermentas, Литва). Секвенирование осуществляли на автоматическом секвенаторе ABI310A (ABI PRISM 310 Genetic Analyzer) в центре коллективного пользования «Геномика» СО РАН (г. Новосибирск). Полученные последовательности проанализировали с помощью пакета программ FASTA (http://www.ebi.ac.uk/fasta33/), на наличие химерных структур проверили программой CHECK CHIMERA (http://rdp8.cme.msu.edu/cgis/chimera) и зарегистрировали в международной EMBL-базе данных под номерами: FR799652, FR799653, FR799680, FR799684, HE565982–HE566014, HE566026–HE566038, HE566041–HE566044, HE584724–HE584729, HE589594–HE589602.
Результаты исследования и их обсуждение
Кишечная микробиота рыб, включающая аэробных, факультативно-анаэробных и облигатных анаэробных бактерий, может быть классифицирована как автохтонная, или собственная (микроорганизмы, колонизирующие эпителий кишечника хозяина), и аллохтонная, или транзиторная. Состав и структура микробного сообщества зависят от экологических условий местообитания и пищевых стратегий рыб.
Сравнительный анализ микробного разнообразия на уровне крупных таксонов: домены Bacteria (EUB) и Archaea (ARC); филы Actinobacteria (ACT), Bacteroidetes (CF), Planctomycetes (PLA), Firmicutes (BLS), Cyanobacteria (CYA) и Spirochaetes (SPI), а также классы филы Proteobacteria: Alpha (ALF), Beta (BET) и Gamma (GAM) с помощью групп-специфичной амплификации не выявил отличий в структуре микробных сообществ кишечной микробиоты омуля трех МЭГ в весенний период. Типичная электрофореграмма приведена на рисунке А. В осенний период у омуля пелагической МЭГ сохраняется такой же широкий спектр таксонов, в то время как у омуля придонно-глубоководной МЭГ разнообразие ниже; не были детектированы представители фил Spirochaetes и Bacteroidetes (рисунок, Б). Таксономический состав микробиоты омуля прибрежной МЭГ осенью сохраняется с перераспределением доминант: значительно уменьшается представленность всех классов протеобактерий, при этом увеличивается количество Spirochaetes (рисунок, В).
Спектр таксономических групп кишечной микробиоты черного байкальского хариуса совпадает с таковым у прибрежного омуля в осенний период, дополнительно у хариуса детектируется байкальский генотип Mycoplasma (рисунок, Г).
Присутствие спирохет и микоплазм в кишечнике рыб может зависеть от типа питания рыб в целом. Микоплазмы и спирохеты отмечены у моллюсков [13]. В пищевом спектре черного байкальского хариуса моллюски являются одним из постоянных компонентов пищи [5]. В питании байкальского омуля моллюски встречаются реже, в основном у рыб, принадлежащих к прибрежной МЭГ.
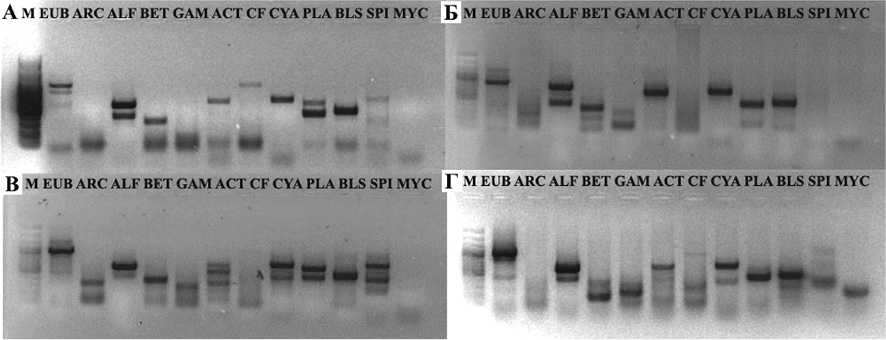
Электрофореграмма ампликонов с суммарной ДНК, выделенной из кишечника байкальского омуля в весенний (А) и осенний (Б, В) периоды, и черного байкальского хариуса (Г) на групп-специфичных праймерах: EUB – домен Bacteria (900 п.н.); ARC – домен Archaea (600 п.н.); ALF – класс Alphaproteobacteria (400 п.н.); BET – класс Betaproteobacteria (350 п.н.); GAM – класс Gammaproteobacteria (150 п.н.); ACT – фила Actinobacteria (640 п.н.); CF – фила Bacteroidetes (1050 п.н.); CYA – фила Cyanobacteria (650 п.н.); PLA – фила Planctomycetes (420 п.н.); BLS – фила Firmicutes (400 п.н.); SPI – фила Spirochaetes (350 п.н.); Мус – байкальский генотип Mycoplasma (150 п.н.); M – маркер молекулярного веса
Микроорганизмы в ассоциациях, в том числе в составе кишечной микробиоты, формируют устойчивые сообщества, способные существовать неопределенно долго за счет поступающей извне энергии и биогенных элементов. Переход макроорганизма на другой тип питания сопровождается появлением новых пищевых компонентов и, как следствие, изменением разнообразия микроорганизмов, это подтверждается результатами анализа микробиоты КТ осенних популяций омуля прибрежной и придонно-глубоководной МЭГ. Пищевые спектры рыб достаточно широки, но качественный состав кормовых объектов, составляющих основу их питания, различен. Различия в составе микробных ассоциаций также отмечены и на уровне таксонов более низкого уровня – родовой принадлежности отдельных представителей как аллохтонной, так и автохтонной микробиоты (таблица). Для черного байкальского хариуса среди представителей классов Gamma и Betaproteobacteria определены последовательности, имеющие высокую гомологию с таковыми из кишечной микробиоты байкальского омуля.
Сравнительный анализ кишечных микробиоценозов черного байкальского хариуса и байкальского омуля
|
Черный байкальский хариус |
Байкальский омуль |
|
Аллохтонные микроорганизмы |
|
|
Azospirillum, Bradyrhizobium, Comamonas, Collimonas, Pseudomonas, Stenotrophomonas |
Pseudoalteromonas, Pseudomonas, Comamonas, Quatrionicoccus, Stenotrophomonas, некультивируемые бактерии сем. Chitinophagaceae и Rubrobacteraceae |
|
Автохтонный микробиоценоз |
|
|
Acinetobacter, Aeromonas, Brevinema, Caulobacter, Clostridium, Deefgea, Delftia, Enterobacter, Escherichia, Flavobacterium, Mycoplasma, Plesiomonas, Pseudomonas, Salmonella, Spironema, Spironucleus barkhanus, Sphingomonas, некультивируемая Deltaproteobacteria |
Enterobacter, Pseudomonas, Salmonella |
Наличие представителей родов Comamonas, Stenotrophomonas и Pseudomonas (Pseudomonas sp., P. putida, P. stutzeri, P. reactans и P. rhodesiae) может быть связано с их присутствием в байкальской воде или в ассоциациях с общими для черного байкальского хариуса и байкальского омуля кормовыми объектами. Также к аллохтонной микробиоте следует отнести некоторых представителей класса Alphaproteobacteria, детектированных в кишечнике черного байкальского хариуса: их ближайшие родственники выделены из ассоциаций с растениями (Bradyrhizobium sp.) и из пресных вод (Azospirillum sp.).
В то же время в составе кишечного микробиоценоза у черного байкальского хариуса определены ближайшие родственники Caulobacter sp. и Sphingomonas sp. (класс Alphaproteobacteria), которые имеют высокую гомологию с некультивируемыми бактериями, выделенными из кишечной микробиоты различных организмов. Так, среди их ближайших гомологов следует отметить некультивируемые бактерии, идентифицированные в кишечнике амура (FR853581), Tamarixia radiata (EF433789) и Sphenophorus levis (FJ626264), симбионтов Nilaparvata lugens (FJ774970), китайского краба Eriocheir sinensis (DQ856499), анчоуса (DQ860013) и антарктических рыб [12]. Несомненно, данные микроорганизмы можно отнести к автохтонной микробиоте байкальских рыб. Представитель рода Delftia (HM587796) (класс Betaproteobacteria, сем. Comamonadaceae) идентифицирован в кишечнике термитов, другие представители этого рода, D. tsuruhatensis и D. lacustris, способны разрушать пептидогликан и проявляют хитиназную активность [8]. К собственной кишечной микробиоте черного байкальского хариуса следует отнести представителей родов Enterobacter и Salmonella, последовательности которых идентичны таковым из кишечника байкальского омуля, а также Escherichia (класс Gammaproteobacteria, сем. Enterobacteriaceae), имеющие высокую гомологию (99 %) с микроорганизмами, культивированными из кишечника насекомых (HQ407235, HQ154567) и полученными в результате молекулярно-генетического анализа слизистой кишечника радуж ной форели [9].
Заключение
Таким образом, исследование структуры микробных сообществ кишечника байкальского омуля трех МЭГ и черного байкальского хариуса с помощью групп-специфичной ПЦР позволило определить спектр таксонов микроорганизмов, населяющих кишечник рыб, и его сезонные изменения. При смене спектра питания увеличивается разнообразие доминирующих микроорганизмов (омуль прибрежной МЭГ), а стабильность питания ведет к формированию устойчивого микробного сообщества с уменьшением числа таксономических единиц (омуль придонно-глубоководной МЭГ). Выявлены представители аллохтонной и автохтонной микробиоты, общие и отличающиеся для разных видов рыб.
Работа выполнена в рамках проекта VI.50.1.4 «Молекулярная экология и эволюция живых систем Центральной Азии на примере рыб, губок и ассоциированной с ними микрофлоры», № гос. рег. 01201353444.
Рецензенты:Анганова Е.В., д.б.н., старший научный сотрудник лаборатории эпидемически и социально значимых инфекций, ФГБНУ НЦ «Проблем здоровья семьи и репродукции человека» ФАНО, г. Иркутск;
Савилов Е.Д., д.м.н., профессор, главный научный сотрудник ФГБНУ НЦ «Проблем здоровья семьи и репродукции человека» ФАНО, г. Иркутск.
Работа поступила в редакцию 26.11.2014.
Библиографическая ссылка
Дзюба Е.В., Суханова Е.В., Деникина Н.Н., Белькова Н.Л. СРАВНИТЕЛЬНЫЙ АНАЛИЗ МИКРОБИОЦЕНОЗОВ КИШЕЧНИКА ЛОСОСЕВИДНЫХ РЫБ С РАЗНЫМИ ПИЩЕВЫМИ СТРАТЕГИЯМИ // Фундаментальные исследования. 2014. № 11-11. С. 2429-2433;URL: https://fundamental-research.ru/ru/article/view?id=35960 (дата обращения: 24.07.2026).