



Известно, что модуляция иммунного ответа организма хорионическим гонадотропином (ХГ) способствует сохранению беременности [10, 13, 14]. При трофобластической болезни на фоне повышенных концентраций ХГ иммунный противоопухолевый ответ снижается, что способствует гематогенному метастазированию опухоли. При наружном генитальном эндометриозе в периферической крови уменьшается количество CD8-позитивных лимфоцитов (маркер цитотоксических T-лимфоцитов), а количество моноцитов возрастает [5]. Этапы внутритимусной дифференцировки клеток от мигрировавшего в орган костномозгового предшественника (пре-Т-клеток) до зрелого Т-лимфоцита, покидающего тимус, связаны с изменением экспрессии фенотипических Т-клеточных маркеров. Положительная селекция в коре определяет дальнейшую дифференцировку на субпопуляцию CD4 и CD8-тимоцитов, которая осуществляется в основном в переходной области – кортико-медуллярном соединении под влиянием взаимодействия с дендритными клетками (ДК) и макрофагами. Здесь же проходит и отрицательная селекция тимоцитов. В результате прошедших дифференцировочных событий в медуллярной области накапливаются клетки с фенотипом CD4+CD8- (Т-хелперы) и CD4- CD8+ (Т-киллеры) [2].
Популяция CD8+-дендритных клеток неоднородна [8, 6]. При различных физиологических и патологических состояниях реакция CD8+-дендритных клеток различна [10]. CD8+-дендритные клетки стимулируют вторичные CD4-позитивные лимфоциты [9]. CD8+-дендритные клетки фагоцитируют антигены и образуют комплекс с пептидами HLA-II [7]. Клетки-предшественники Т-лимфоцитов (CD4+, CD8+) синтезируют и содержат серотонин [11].
Методами люминесцентной гистохимии в опытах на белых лабораторных крысах при их беременности установлено, что интенсивность свечения моноаминов в дендритных макрофагах премедуллярной зоны тимусной дольки значительно увеличивается [3]. Обнаружено, что концентрация иммуносупрессорного моноамина серотонина преобладает в тимоцитах мозгового вещества долек тимуса [4].
Цель исследования: изучение популяции CD8-позитивных клеток в различных морфофункциональных зонах долек тимуса мышей-самок у интактных, контрольных и опытных с введением хорионического гонадотропина.
Материалы и методы исследования
Объектом исследования служил тимус 40 белых лабораторных мышей-самок массой 30 г. Животные были разделены на 3 группы: I – интактная группа (n = 10); II – контрольная (n = 10), которым внутримышечно вводили 0,02 мл физраствора; III – опытная с внутримышечным введением 0,02 мл раствора ХГ: III А – введение ХГ в течение 1 недели (2 раза через каждые 3 дня, n = 5), III Б – введение ХГ в течение 2 недель (4 раза через каждые 3 дня, n = 5), III В – введение ХГ в течение 3 недель (6 раз через каждые 3 дня, n = 5), III Г – введение ХГ в течение 4 недель (8 раз через каждые 3 дня, n = 5). Введение препаратов проводилось с соблюдением правил асептики и антисептики. Все процедуры по уходу за мышами осуществляли согласно нормам и правилам обращения с лабораторными животными. Тимус извлекали под наркозом после завершения экспериментального воздействия.
Срезы тимуса обрабатывались на CD8+-клеток по стандартной методике: сначала подавлялась пероксидазная активность путем инкубации срезов в 3 % растворе Н2О2 в течение 30 минут с последующей промывкой 0,1М фосфатным буфером. Для блока неспецифического связывания срезы инкубировались в течение 1 часа в 10 % козьей сыворотке, после чего к ним были добавлены первичные антитела к белкам CD8 на 18 часов при температуре 4 °С. В качестве вторых антител были использованы антивидовые антииммуноглобулиновые биотилированные антитела. С целью выявления биотиновой метки срезы обрабатывались авидин-пероксидазным комплексом. Метод основан на высоком сродстве авидина к биотину. Пероксидазную активность проявляли в инкубационной среде с диаминобензидином. В результате ферментативной реакции субстрат превращался в нерастворимый продукт коричневого цвета, совпадающий по локализации с местонахождением белка [12]. Препараты анализировались под световым микроскопом Микмед-5. В качестве общеморфологического метода использовали окраску срезов тимуса гематоксилин-эозином. Производили подсчет и морфометрический анализ CD8-позитивных клеток в 10 полях зрения с вычислением среднего значения показателей, среднеквадратичного отклонения и критерия достоверности Р < 0,05. Выводы опираются на достоверные результаты с использованием U-критерия Вилкоксона – Манна – Уитни. Морфометрический анализ включал измерение размеров коркового и мозгового веществ долек тимуса, которое проводилось после получения фотографий при увеличении х100 (объектив 10 и окуляр 10), а определение размеров клеточных структур при увеличении х400, х600 и х1000 (объектив 40, 60, 100 и окуляр 10) с использованием светового микроскопа МИКМЕД-5. Для морфометрической оценки использовалась программа «Sigma Scan Pro 5.0». Количество клеток измерялось в десяти квадратах площадью 0,04 мм2 в сравниваемых морфофункциональных зонах долек тимуса (корковое, кортико-медуллярная и мозговое вещество).
Результаты исследования и их обсуждение
Изучение распределения CD8-позитивных клеток в дольках тимуса интактных животных показало, что наибольшее количество их выявляется в мозговом веществе (2,0 ± 0,2) и кортикомедуллярной зоне (1,4 ± 0,09) (рис. 1). В корковом веществе долек тимуса визуализируется небольшое количество CD8-позитивных клеток – 0,6 ± 0,05 (табл. 1).
CD8-позитивные клетки (размер 8–12 мкм) имеют разнообразную форму (овальную, округлую, сферическую) и лежат в виде вкраплений среди тимоцитов и эпителиальных клеток тимуса. Коричневый цвет в клетках совпадает по локализации с местонахождением белка CD8+. Площадь клеток в среднем составляет 91,12 ± 12,00 мкм2 (табл. 2).
Анализ результатов экспериментального исследования показал, что введение хорионического гонадотропина приводит к изменению количества CD8-позитивных клеток в дольках тимуса в сравнении с контрольной группой. Исследуемые клетки вилочковой железы в основном выявлялись в корковом, мозговом веществах и кортико-медуллярной зоне долек. В корковом и мозговом веществах долек железы CD8-позитивные лимфоциты располагались диффузно. CD8-позитивные клеточные структуры кортико-медуллярной зоны по морфологическим свойствам и расположению совпадают с люминесцирующими гранулярными клетками, идентифицированные как дендритные макрофаги [3]. Известно, что дендритные макрофаги имеют высокие уровни экспрессии CD8+ пептидов [1].
По мнению Kronin V., Fitzmaurice C.J. et. al. [9], Mason D.Y., Cordel J.L. et. al. [12] CD8+-белок экспрессируется Т-цитотоксическими клетками иммунной системы и дендритными макрофагами. В функции CD8+-дендритных клеток входит стимуляция вторичных CD4+-лимфоцитов, которая зависит от интерлейкина-2 и интерлейкина-4.
Наши исследования показали, что введение ХГ в течение 1 недели приводит к росту количества CD8-позитивных клеток в мозговом веществе долек тимуса в 4,4 раза (8,8 ± 0,9), в корковом веществе – в 4 раза (2,4 ± 0,4), тогда как в кортикомедуллярной зоне в 2,3 раза снижается количество CD8-позитивных клеток.
Таблица 1
Количественное распределение CD8-позитивных клеток в морфофункциональных зонах долек тимуса мышей: интактных, контрольных (с введением физ. раствора) и при введении хорионического гонадотропина (M ± m)
|
Морфофункциональные зоны долек тимуса |
Группа экспериментальных мышей |
||||||||
|
Сроки опытов |
1 неделя |
2 неделя |
3 неделя |
4 неделя |
|||||
|
интактные |
ХГ |
Физ. р-р |
ХГ |
Физ. р-р |
ХГ |
Физ. р-р |
ХГ |
Физ. р-р |
|
|
Корковое вещество |
0,6 ± 0,05 |
2,4 ± 0,4 |
1,2 ± 0,06 |
3,5 ± 0,5 |
– |
5,2 ± 0,6 |
1,4 ± 0,06 |
8,0 ± 0,4* |
1,4 ± 0,06 |
|
Кортикомедуллярная зона |
1,4 ± 0,09 |
0,6 ± 0,05 |
1,0 ± 0,05 |
2,8 ± 0,2 |
1,0 ± 0,05 |
3,6 ± 0,5 |
1,4 ± 0,06 |
3,4 ± 0,05* |
1,4 ± 0,06 |
|
Мозговое вещество |
2,0 ± 0,2 |
8,8 ± 0,9 |
2,2 ± 0,06 |
7,9 ± 0,6 |
1,9 ± 0,05 |
6,0 ± 0,3 |
2,0 ± 0,05 |
7,6 ± 0,03* |
2,0 ± 0,05 |
Примечание. *р < 0,05 по сравнению с интактной группой.
Таблица 2
Морфометрические показатели CD8-позитивных клеток в тимусе мышей: интактных, контрольных (с введением физ. раствора) и при введении хорионического гонадотропина, мкм2 (М ± m)
|
Площадь клеток |
Группа экспериментальных мышей |
|||||
|
интактные |
Физ. р-р |
ХГ 1 неделя |
ХГ 2 недели |
ХГ 3 недели |
ХГ 4 недели |
|
|
91,12 ± 12,00 |
45,99 ± 11,66 |
103,35 ± 18,61 |
75,77 ± 20,93 |
127,28 ± 17,08 |
191 ± 15,34 |
|
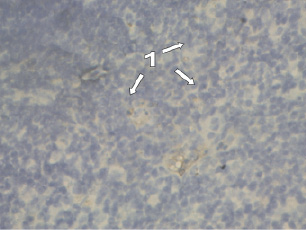
Рис. 1. Тимус мыши. Интактная группа. Иммуногистохимическая реакция с антителами СD8+. Микроскоп Микмед-5. Об. 40, ок. 15. 1 – CD8+-позитивные клетки
В последующие сроки (2, 3 недели) введения ХГ во всех исследуемых зонах тимуса количество CD8-позитивных клеток продолжает увеличиваться по сравнению с интактными животными (табл. 1).
Через 4 недели экспериментального воздействия гормона количественные показатели содержания цитотоксических лимфоцитов в дольках тимуса остаются высокими.
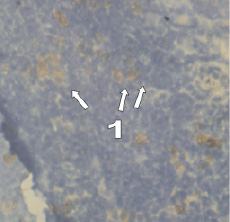
Рис. 2. Тимус мыши. Внутримышечное введение хорионического гонадотропина в течение 4 недель. Иммуногистохимическая реакция с антителами СD8+. Микроскоп Микмед-5. Об. 40, ок. 15. 1 – CD8+-позитивные клетки
Морфометрический анализ CD8-позитивных клеток тимуса на фоне введения ХГ показал увеличение площади исследуемых клеток по сравнению с интактными животными (у интактных животных – 91,12 ± 12,00 мкм2; через 1 неделю введения ХГ – 103,35 ± 18,611 мкм2; через 3 недели – 127,28 ± 17,08 мкм2; через 4 недели – 191 ± 15,34 мкм2) (табл. 2).
В серии работ [10, 13, 14] было показано, что хорионический гонадотропин является основным иммунодепрессантом в организме. Наши исследования показали, что под влиянием ХГ происходит увеличение количества CD8-позитивных клеток у мышей-самок в центральном органе иммунитета и увеличению площади CD8+-позитивных клеток, что свидетельствует об усилении функциональной активности этих клеток.
Дендритные клетки также экспрессируют CD8 + белок, продуцируют интерферон, стимулируют воспалительные процессы, обеспечивают толерантность к аутогенным клеткам. В наших исследованиях обращает на себя внимание ситуация с дендритными макрофагами кортикомедуллярной зоны вилочковой железы, где количество CD8+-позитивных дендритных клеток уменьшается через 1 неделю введения ХГ, а в последующие сроки – через 2, 3, 4 недели введения ХГ ‒ значительно увеличивается их количество и площадь клеток.
В мозговом веществе долек через неделю введения ХГ достоверно увеличивается количество CD8-позитивных клеток по сравнению с интактной группой, очевидно, хорионический гонадотропин способствует усилению образования цитотоксических лимфоцитов в тимусе. Через 2, 3 и 4 недели введения ХГ отмечается незначительное уменьшение CD8 + клеток в сравнении с 1-недельным сроком воздействия. Возможно, усилена миграция цитотоксических лимфоцитов из мозгового вещества долек тимуса.
Заключение
Таким образом, введение хорионического гонадотропина в течение 1, 2, 3 и 4 недель оказывает иммуномодулирующее влияние на микроструктуры тимуса мышей-самок и приводит:
1) к увеличению количества CD8+-клеток в корковом и мозговом веществе, кортикомедуллярной зоне долек тимуса;
2) к увеличению площади CD8+ позитивных клеток, что свидетельствует об усилении функциональной активности этих клеток.
Работа выполнена на кафедре медицинской биологии с курсом микробиологии и вирусологии. Госбюджетная тема 0120.085.1887 от 2008 г. «Гистохимия биогенных аминов в морфофункциональном состоянии органов и тканей в норме и эксперименте».
Рецензенты:
Алексеев В.В., д.б.н., профессор кафедры биоэкологии и географии, декан факультета естественнонаучного образования, ФГБОУ ВПО «Чувашский государственный педагогический университет им. И.Я. Яковлева», г. Чебоксары;
Долгов И.Ю., д.м.н., профессор, главный врач БУ ЧР «Республиканский клинический онкологический диспансер», г. Чебоксары.
Работа поступила в редакцию 23.07.2014.
Библиографическая ссылка
Ялалетдинова Л.Р., Ястребова С.А., Сергеева В.Е. ЭФФЕКТ ХОРИОНИЧЕСКОГО ГОНАДОТРОПИНА НА CD8-ПОЗИТИВНЫЕ СТРУКТУРЫ ВИЛОЧКОВОЙ ЖЕЛЕЗЫ // Фундаментальные исследования. 2014. № 7-5. С. 1075-1079;URL: https://fundamental-research.ru/ru/article/view?id=34832 (дата обращения: 15.07.2026).